Hypermigration of macrophages through the concerted action of GRA effectors on NF-κB/p38 signaling and host chromatin accessibility potentiates Toxoplasma dissemination
- PMID: 39207098
- PMCID: PMC11481493
- DOI: 10.1128/mbio.02140-24
Hypermigration of macrophages through the concerted action of GRA effectors on NF-κB/p38 signaling and host chromatin accessibility potentiates Toxoplasma dissemination
Abstract
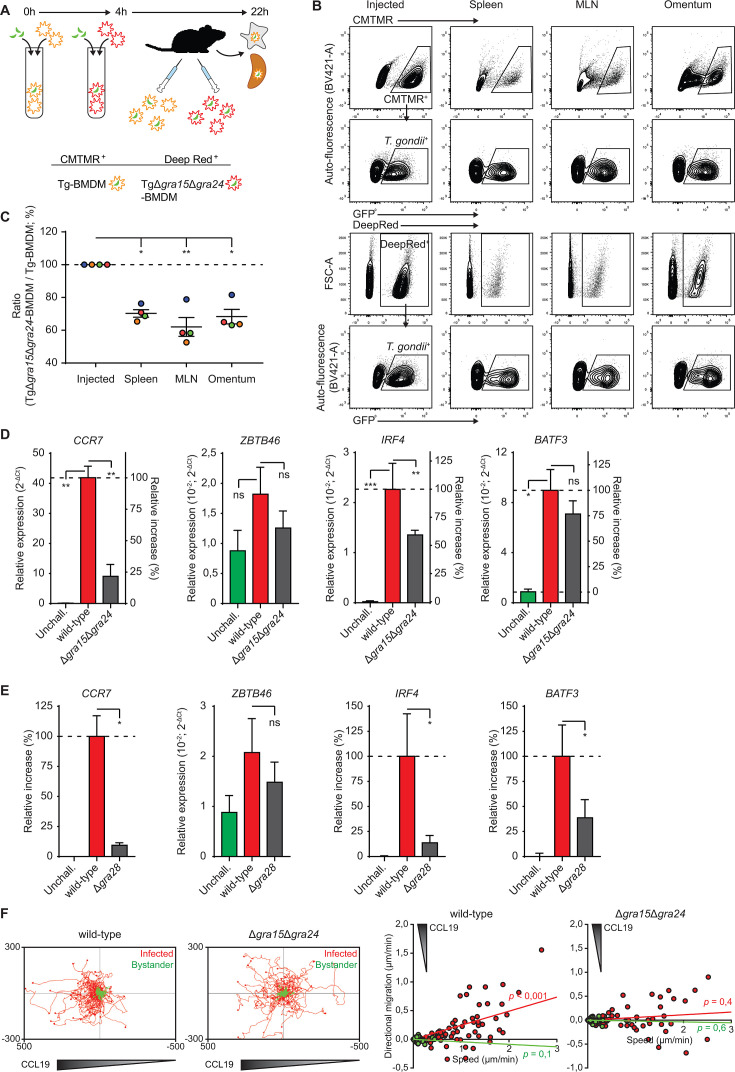
Mononuclear phagocytes facilitate the dissemination of the obligate intracellular parasite Toxoplasma gondii. Here, we report how a set of secreted parasite effector proteins from dense granule organelles (GRA) orchestrates dendritic cell-like chemotactic and pro-inflammatory activation of parasitized macrophages. These effects enabled efficient dissemination of the type II T. gondii lineage, a highly prevalent genotype in humans. We identify novel functions for effectors GRA15 and GRA24 in promoting CCR7-mediated macrophage chemotaxis by acting on NF-κB and p38 mitogen-activated protein kinase signaling pathways, respectively, with contributions by GRA16/18 and counter-regulation by effector TEEGR. Furthermore, GRA28 boosted chromatin accessibility and GRA15/24/NF-κB-dependent transcription at the Ccr7 gene locus in primary macrophages. In vivo, adoptively transferred macrophages infected with wild-type T. gondii outcompeted macrophages infected with a GRA15/24 double mutant in migrating to secondary organs in mice. The data show that T. gondii, rather than being passively shuttled, actively promotes its dissemination by inducing a finely regulated pro-migratory state in parasitized human and murine phagocytes via co-operating polymorphic GRA effectors.
Importance: Intracellular pathogens can hijack the cellular functions of infected host cells to their advantage, for example, for intracellular survival and dissemination. However, how microbes orchestrate the hijacking of complex cellular processes, such as host cell migration, remains poorly understood. As such, the common parasite Toxoplasma gondii actively invades the immune cells of humans and other vertebrates and modifies their migratory properties. Here, we show that the concerted action of a number of secreted effector proteins from the parasite, principally GRA15 and GRA24, acts on host cell signaling pathways to activate chemotaxis. Furthermore, the protein effector GRA28 selectively acted on chromatin accessibility in the host cell nucleus to selectively boost host gene expression. The joint activities of GRA effectors culminated in pro-migratory signaling within the infected phagocyte. We provide a molecular framework delineating how T. gondii can orchestrate a complex biological phenotype, such as the migratory activation of phagocytes to boost dissemination.
Keywords: cell signaling pathway; host-pathogen; immune cell migration; intracellular parasitism; mononuclear phagocyte.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Update of
-
Hypermigration of macrophages through the concerted action of GRA effectors on NF-κB/p38 signaling and host chromatin accessibility potentiates Toxoplasma dissemination.Ten Hoeve AL, Rodriguez ME, Säflund M, Michel V, Magimel L, Ripoll A, Yu T, Hakimi MA, Saeij JPJ, Ozata DM, Barragan A. Ten Hoeve AL, et al. bioRxiv [Preprint]. 2024 Feb 6:2024年02月06日.579146. doi: 10.1101/2024.02.06.579146. bioRxiv. 2024. Update in: mBio. 2024 Oct 16;15(10):e0214024. doi: 10.1128/mbio.02140-24. PMID: 38370679 Free PMC article. Updated. Preprint.
References
-
- Bosurgi L, Cao YG, Cabeza-Cabrerizo M, Tucci A, Hughes LD, Kong Y, Weinstein JS, Licona-Limon P, Schmid ET, Pelorosso F, Gagliani N, Craft JE, Flavell RA, Ghosh S, Rothlin CV. 2017. Macrophage function in tissue repair and remodeling requires IL-4 or IL-13 with apoptotic cells. Science 356:1072–1076. doi:10.1126/science.aai8132 - DOI - PMC - PubMed
-
- Miller JC, Brown BD, Shay T, Gautier EL, Jojic V, Cohain A, Pandey G, Leboeuf M, Elpek KG, Helft J, Hashimoto D, Chow A, Price J, Greter M, Bogunovic M, Bellemare-Pelletier A, Frenette PS, Randolph GJ, Turley SJ, Merad M, Immunological Genome Consortium . 2012. Deciphering the transcriptional network of the dendritic cell lineage. Nat Immunol 13:888–899. doi:10.1038/ni.2370 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials