An in vitro method for inducing titan cells reveals novel features of yeast-to-titan switching in the human fungal pathogen Cryptococcus gattii
- PMID: 35969643
- PMCID: PMC9426920
- DOI: 10.1371/journal.ppat.1010321
An in vitro method for inducing titan cells reveals novel features of yeast-to-titan switching in the human fungal pathogen Cryptococcus gattii
Erratum in
-
Correction: An in vitro method for inducing titan cells reveals novel features of yeast-to-titan switching in the human fungal pathogen Cryptococcus gattii.Saidykhan L, Correia J, Romanyuk A, Peacock AFA, Desanti GE, Taylor-Smith L, Makarova M, Ballou ER, May RC. Saidykhan L, et al. PLoS Pathog. 2022 Nov 29;18(11):e1011001. doi: 10.1371/journal.ppat.1011001. eCollection 2022 Nov. PLoS Pathog. 2022. PMID: 36445880 Free PMC article.
Abstract
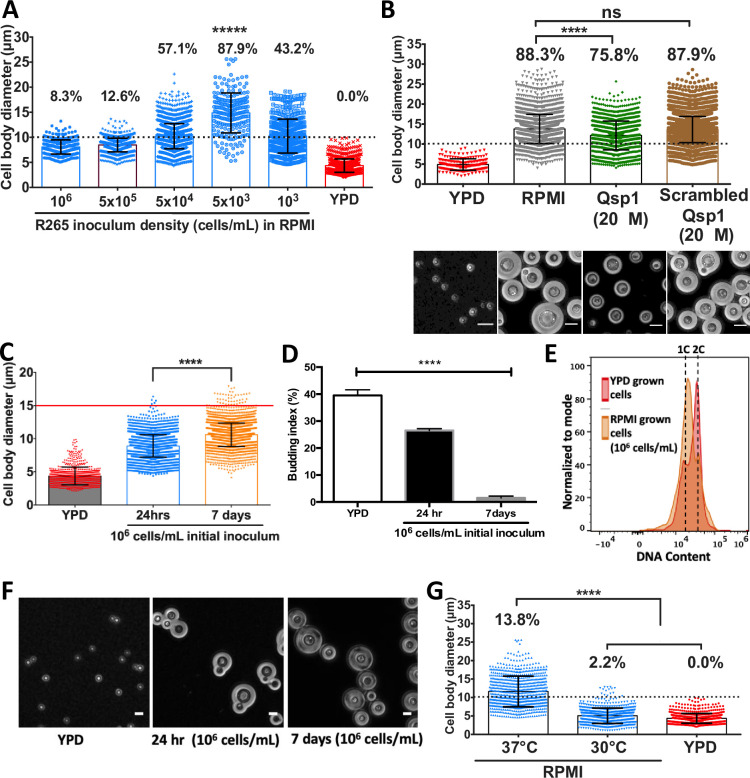
Cryptococcosis is a potentially lethal fungal infection of humans caused by organisms within the Cryptococcus neoformans/gattii species complex. Whilst C. neoformans is a relatively common pathogen of immunocompromised individuals, C. gattii is capable of acting as a primary pathogen of immunocompetent individuals. Within the host, both species undergo morphogenesis to form titan cells: exceptionally large cells that are critical for disease establishment. To date, the induction, defining attributes, and underlying mechanism of titanisation have been mainly characterized in C. neoformans. Here, we report the serendipitous discovery of a simple and robust protocol for in vitro induction of titan cells in C. gattii. Using this in vitro approach, we reveal a remarkably high capacity for titanisation within C. gattii, especially in strains associated with the Pacific Northwest Outbreak, and characterise strain-specific differences within the clade. In particular, this approach demonstrates for the first time that cell size changes, DNA amplification, and budding are not always synchronous during titanisation. Interestingly, however, exhibition of these cell cycle phenotypes was correlated with genes associated with cell cycle progression including CDC11, CLN1, BUB2, and MCM6. Finally, our findings reveal exogenous p-Aminobenzoic acid to be a key inducer of titanisation in this organism. Consequently, this approach offers significant opportunities for future exploration of the underlying mechanism of titanisation in this genus.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
MeSH terms
Substances
Grants and funding
- NC/R001472/1/NC3RS_/National Centre for the Replacement, Refinement and Reduction of Animals in Research/United Kingdom
- BB/R008485/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- 211241/Z/18/Z/WT_/Wellcome Trust/United Kingdom
- BB/M014525/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Research Materials