Pooled CRISPR screening identifies m6A as a positive regulator of macrophage activation
- PMID: 33910903
- PMCID: PMC8081357
- DOI: 10.1126/sciadv.abd4742
Pooled CRISPR screening identifies m6A as a positive regulator of macrophage activation
Abstract
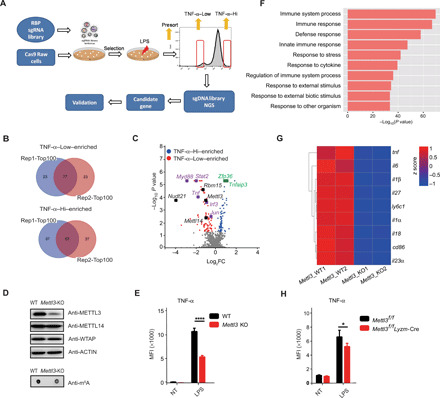
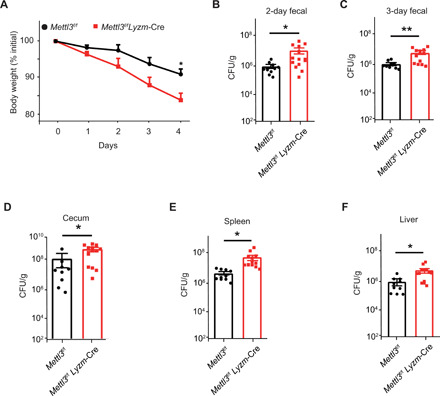
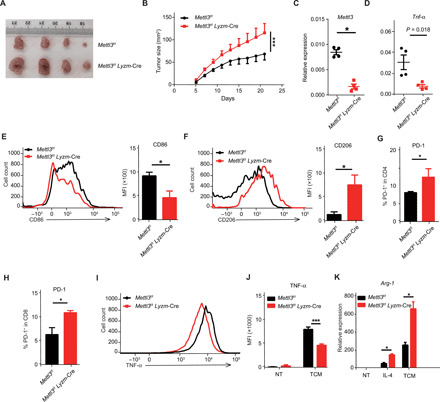
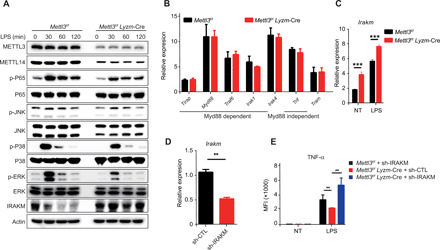
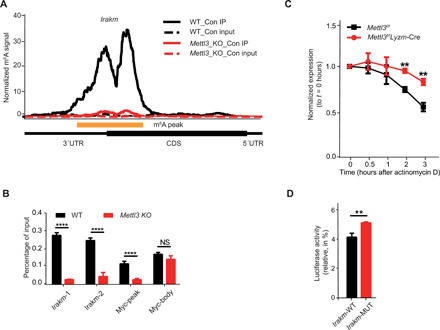
m6A RNA modification is implicated in multiple cellular responses. However, its function in the innate immune cells is poorly understood. Here, we identified major m6A "writers" as the top candidate genes regulating macrophage activation by LPS in an RNA binding protein focused CRISPR screening. We have confirmed that Mettl3-deficient macrophages exhibited reduced TNF-α production upon LPS stimulation in vitro. Consistently, Mettl3 flox/flox;Lyzm-Cre mice displayed increased susceptibility to bacterial infection and showed faster tumor growth. Mechanistically, the transcripts of the Irakm gene encoding a negative regulator of TLR4 signaling were highly decorated by m6A modification. METTL3 deficiency led to the loss of m6A modification on Irakm mRNA and slowed down its degradation, resulting in a higher level of IRAKM, which ultimately suppressed TLR signaling-mediated macrophage activation. Our findings demonstrate a previously unknown role for METTL3-mediated m6A modification in innate immune responses and implicate the m6A machinery as a potential cancer immunotherapy target.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Kawai T., Akira S., Toll-like receptors and their crosstalk with other innate receptors in infection and immunity. Immunity 34, 637–650 (2011). - PubMed
-
- Murray P. J., Macrophage polarization. Annu. Rev. Physiol. 79, 541–566 (2017). - PubMed
-
- Vitale I., Manic G., Coussens L. M., Kroemer G., Galluzzi L., Macrophages and metabolism in the tumor microenvironment. Cell Metab. 30, 36–50 (2019). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases