The role of Toll-like receptor 9 in a murine model of Cryptococcus gattii infection
- PMID: 33446850
- PMCID: PMC7809259
- DOI: 10.1038/s41598-021-80959-5
The role of Toll-like receptor 9 in a murine model of Cryptococcus gattii infection
Abstract
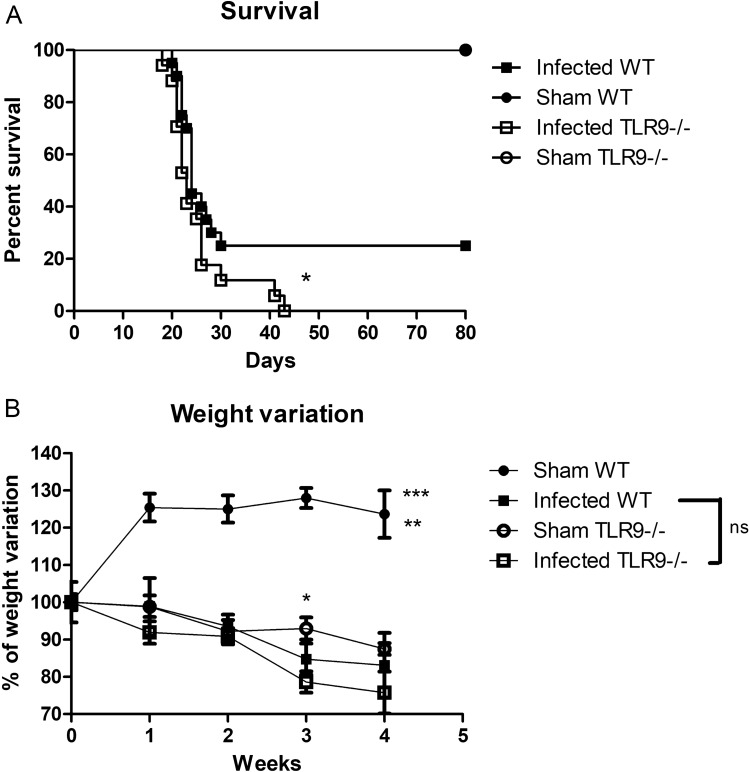
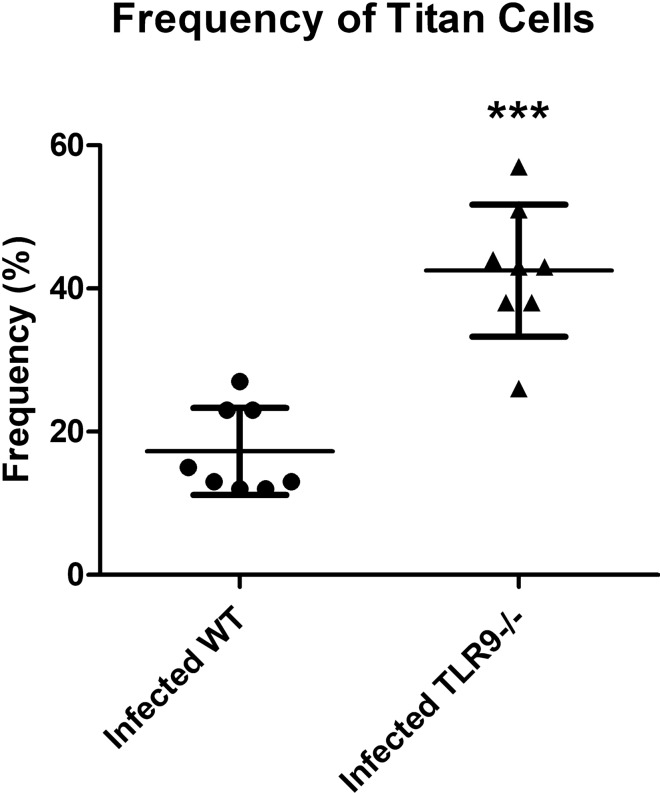
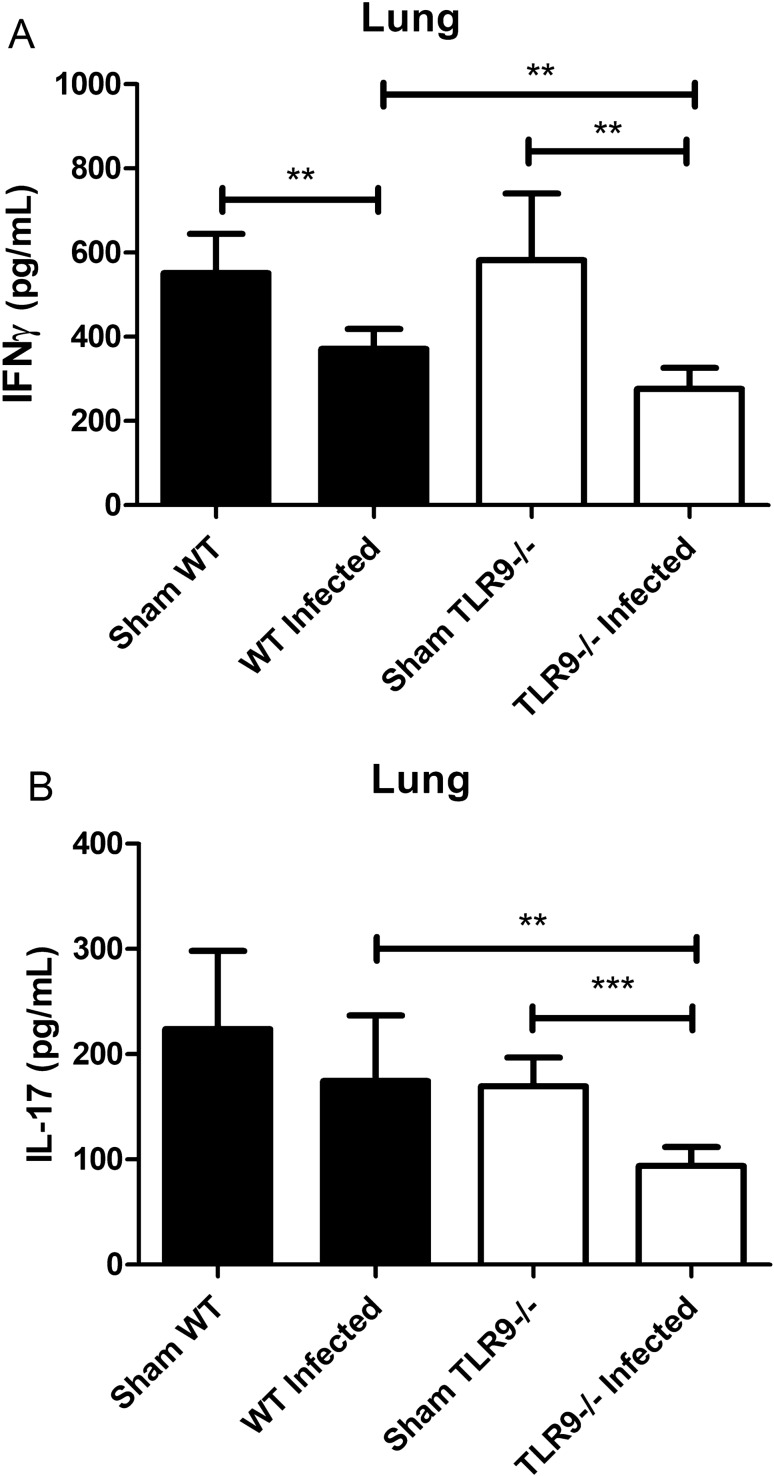
Toll-like receptor 9 (TLR9) is crucial to the host immune response against fungi, such as Candida albicans, Aspergillus fumigatus and Cryptococcus neoformans, but its importance in Cryptococcus gattii infection is unknown. Our study aimed to understand the role of TLR9 during the course of experimental C. gattii infection in vivo, considering that the cryptococcal DNA interaction with the receptor could contribute to host immunity even in an extremely susceptible model. We inoculated C57BL/6 (WT) and TLR9 knock-out (TLR9-/-) mice intratracheally with 104 C. gattii yeast cells. TLR9-/- mice had a higher mortality rate compared to WT mice and more yeast cells that had abnormal size, known as titan cells, in the lungs. TLR9-/- mice also had a greater number of CFUs in the spleen and brain than WT mice, in addition to having lower levels of IFN-γ and IL-17 in the lung. With these markers of aggressive cryptococcosis, we can state that TLR9-/- mice are more susceptible to C. gattii, probably due to a mechanism associated with the decrease of a Th1 and Th17-type immune response that promotes the formation of titan cells in the lungs. Therefore, our results indicate the participation of TLR9 in murine resistance to C. gattii infection.
Conflict of interest statement
The authors declare no competing interests.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases