CRISPR/Cas9 knockout of female-biased genes AeAct-4 or myo-fem in Ae. aegypti results in a flightless phenotype in female, but not male mosquitoes
- PMID: 33338046
- PMCID: PMC7781531
- DOI: 10.1371/journal.pntd.0008971
CRISPR/Cas9 knockout of female-biased genes AeAct-4 or myo-fem in Ae. aegypti results in a flightless phenotype in female, but not male mosquitoes
Abstract
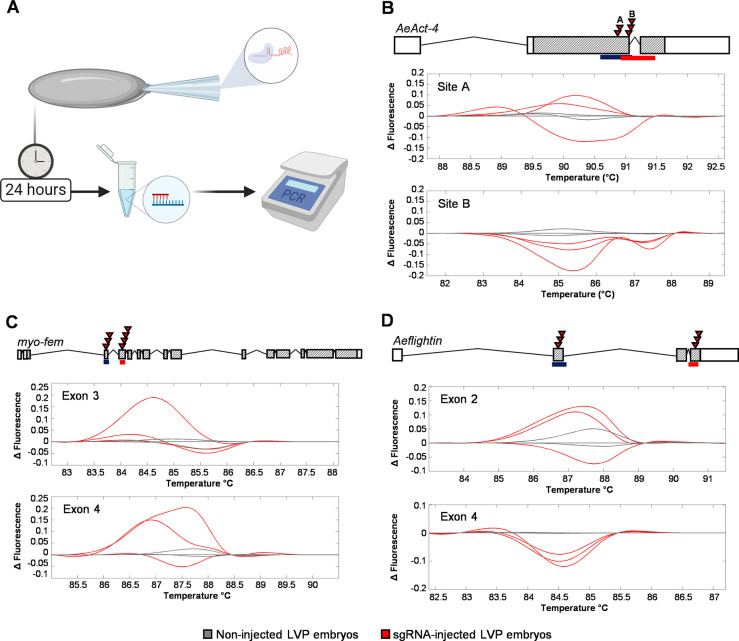
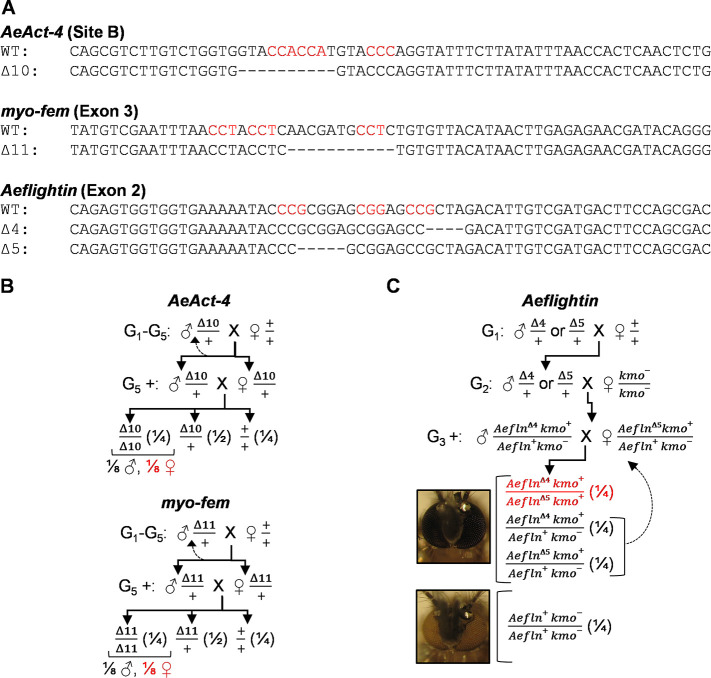
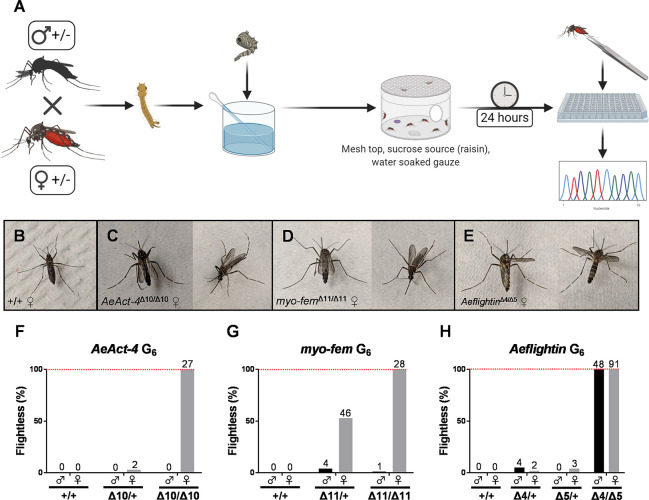
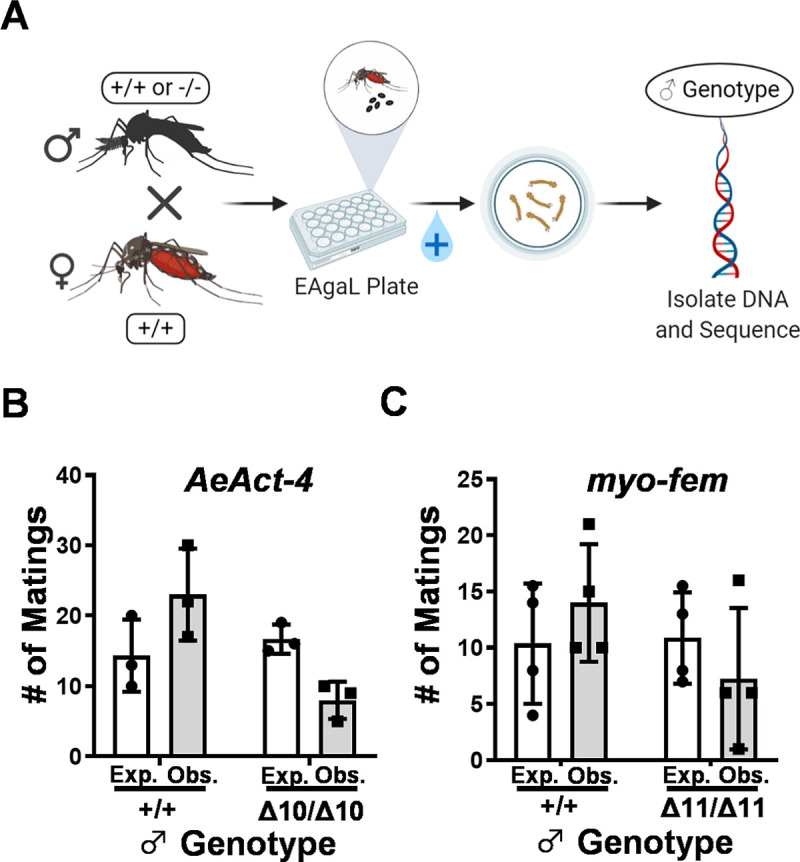
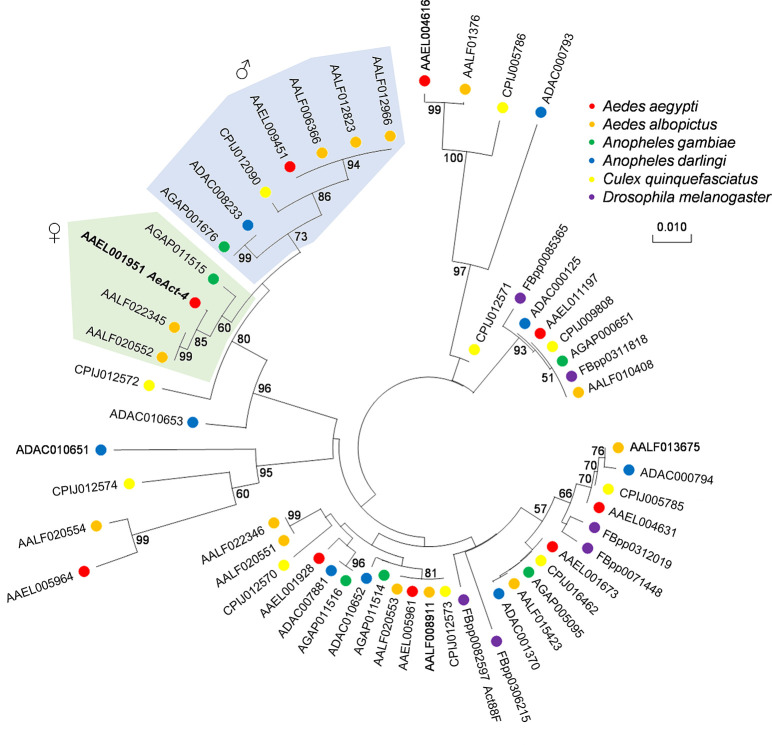
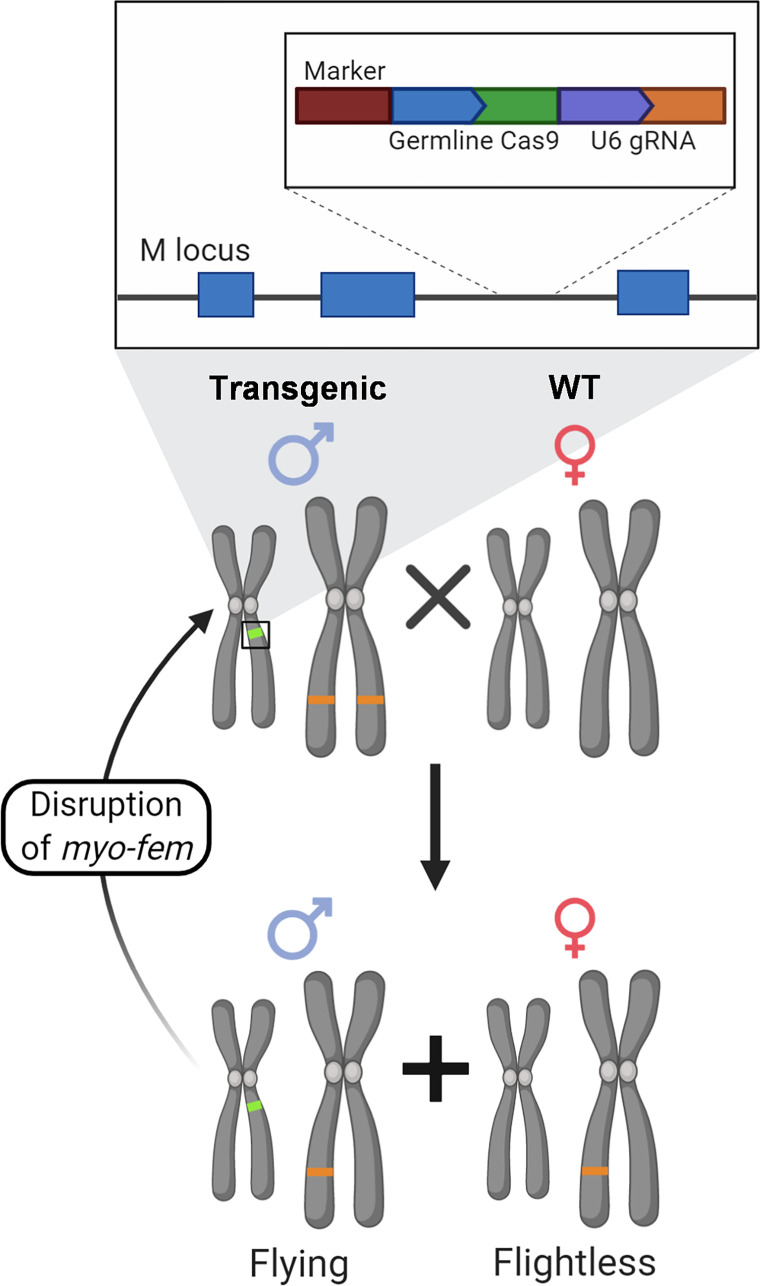
Aedes aegypti is a vector of dengue, chikungunya, and Zika viruses. Current vector control strategies such as community engagement, source reduction, and insecticides have not been sufficient to prevent viral outbreaks. Thus, interest in novel strategies involving genetic engineering is growing. Female mosquitoes rely on flight to mate with males and obtain a bloodmeal from a host. We hypothesized that knockout of genes specifically expressed in female mosquitoes associated with the indirect flight muscles would result in a flightless female mosquito. Using CRISPR-Cas9 we generated loss-of-function mutations in several genes hypothesized to control flight in mosquitoes, including actin (AeAct-4) and myosin (myo-fem) genes expressed specifically in the female flight muscle. Genetic knockout of these genes resulted in 100% flightless females, with homozygous males able to fly, mate, and produce offspring, albeit at a reduced rate when compared to wild type males. Interestingly, we found that while AeAct-4 was haplosufficient, with most heterozygous individuals capable of flight, this was not the case for myo-fem, where about half of individuals carrying only one intact copy could not fly. These findings lay the groundwork for developing novel mechanisms of controlling Ae. aegypti populations, and our results suggest that this mechanism could be applicable to other vector species of mosquito.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- CDC. Surveillance and Control of Aedes aegypti and Aedes albopictus in the United States. 2017.
-
- CDC. Mosquito life cycle. 2018.
-
- Zettel C, Kaufman P. Yellow fever mosquito Aedes aegypti (Linnaeus) (Insecta: Diptera: Culicidae). Featured Creatures [Internet]. 2009:[8 p.].
-
- CDC. Help Control Mosquitoes that Spread Dengue, Chikungunya, and Zika Viruses. 2015.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical