Phagosomal F-Actin Retention by Cryptococcus gattii Induces Dendritic Cell Immunoparalysis
- PMID: 33234684
- PMCID: PMC7701985
- DOI: 10.1128/mBio.01821-20
Phagosomal F-Actin Retention by Cryptococcus gattii Induces Dendritic Cell Immunoparalysis
Abstract
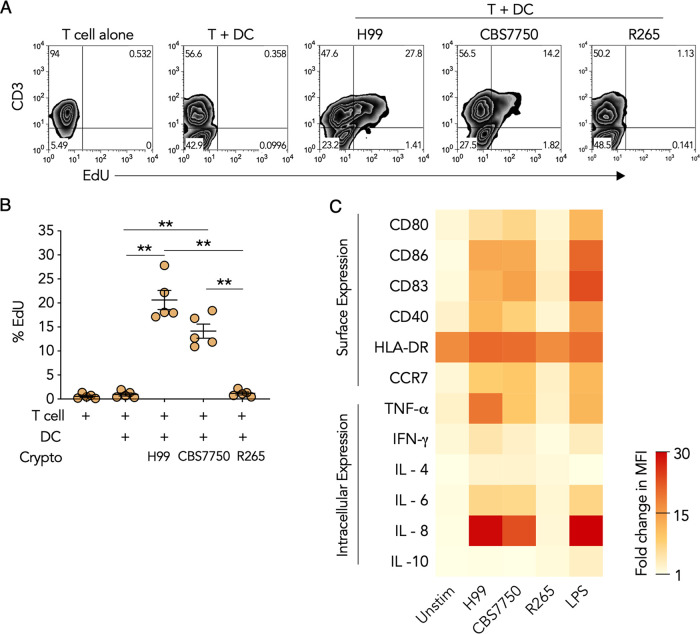
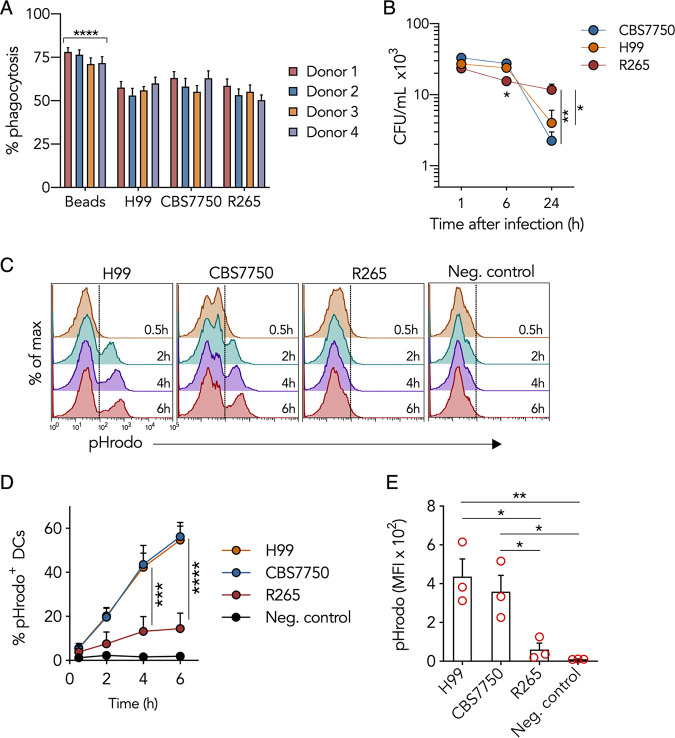
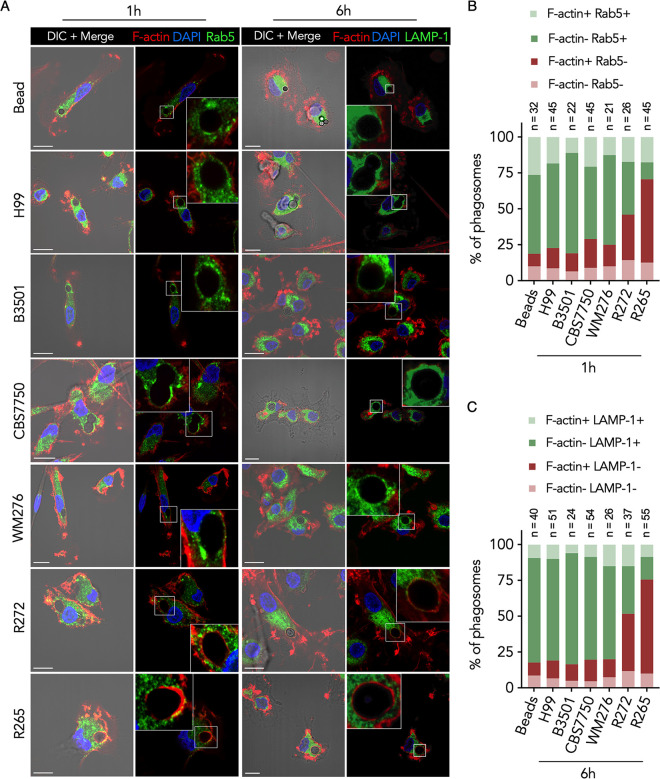
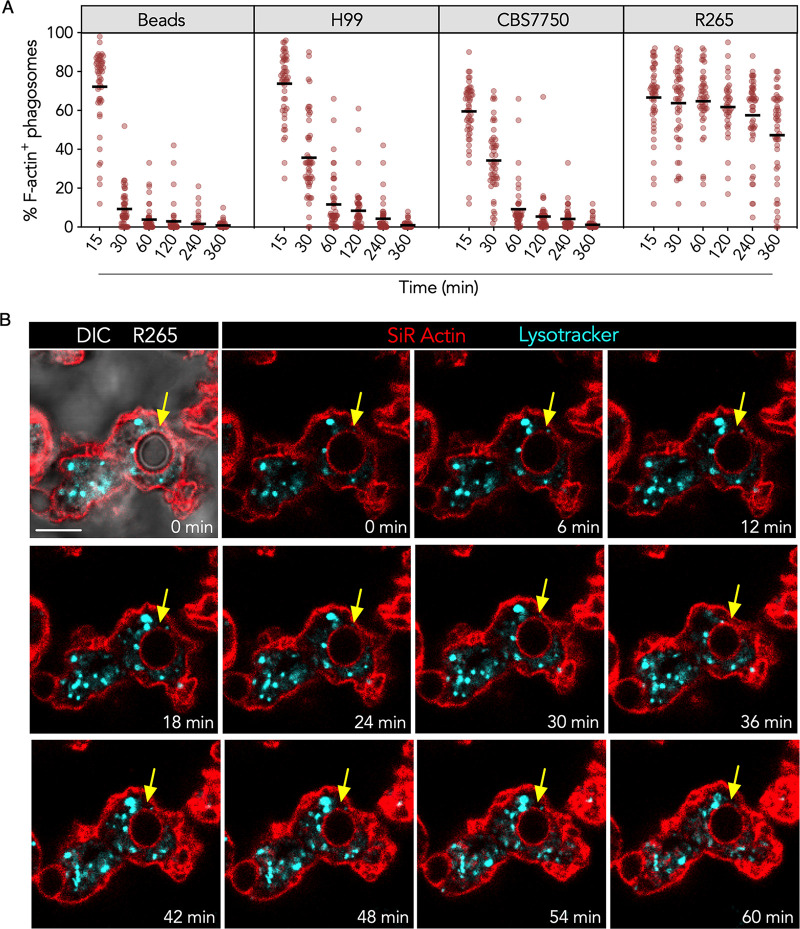
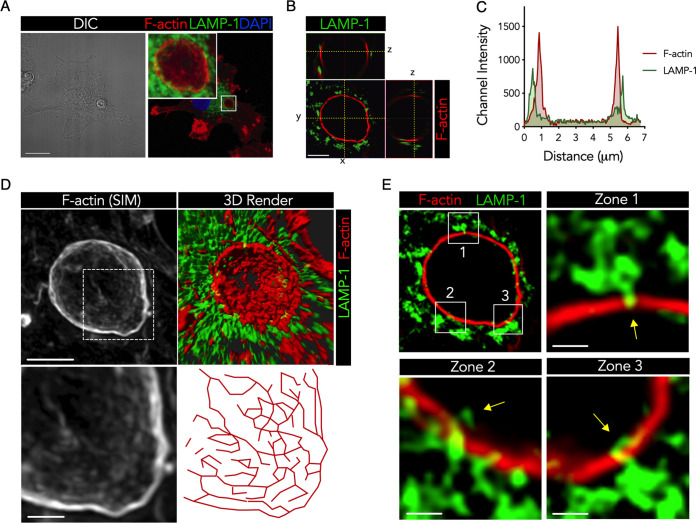
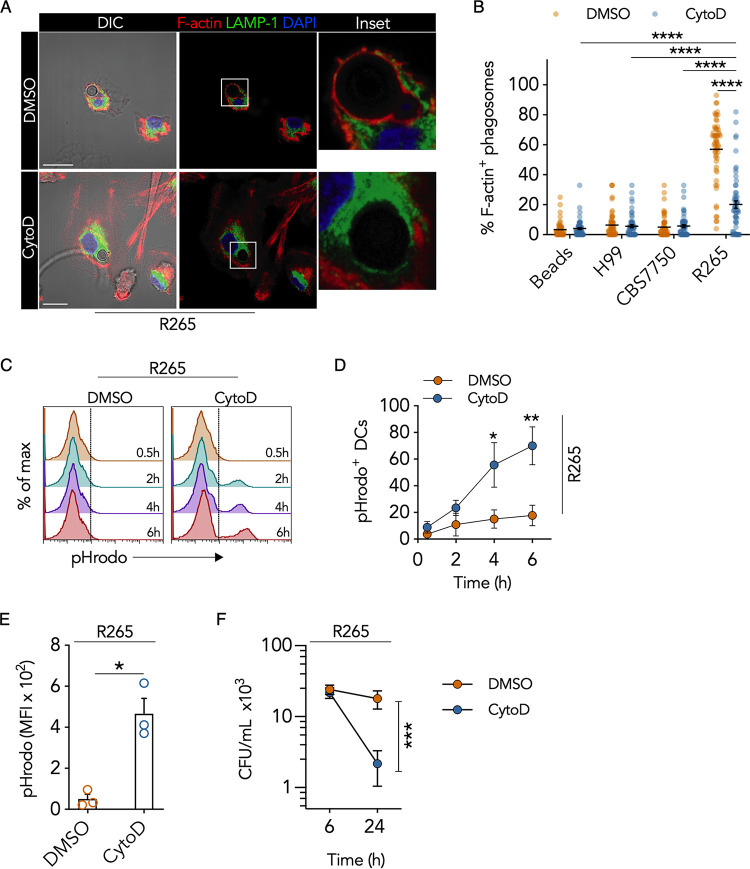
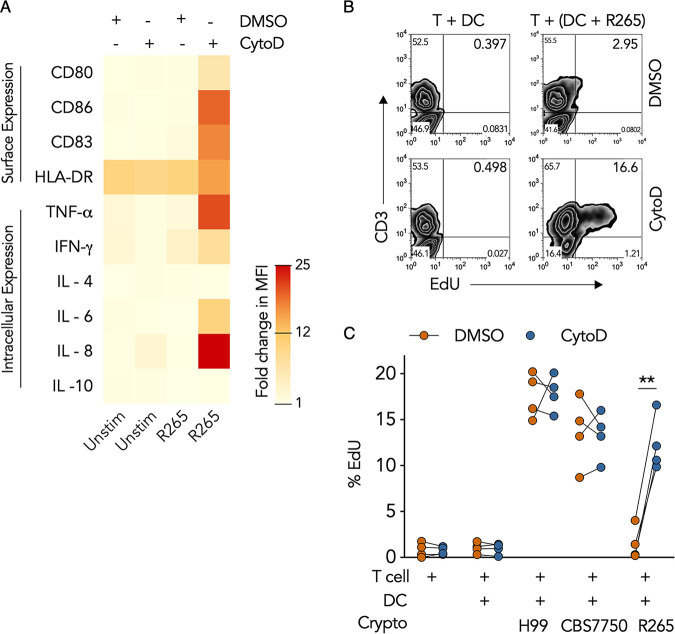
Cryptococcus gattii is a major cause of life-threatening mycosis in immunocompetent individuals and responsible for the ongoing epidemic outbreak of cryptococcosis in the Pacific Northwest of North America. This deadly fungus is known to evade important host immune responses, including dendritic cell (DC) maturation and concomitant T cell immunity, via immune evasion mechanisms that remain unclear. Here, we demonstrate that primary human DCs phagocytose C. gattii but the maturation of phagosomes to phagolysosomes was blocked as a result of sustained filamentous actin (F-actin) that entrapped and concealed the phagosomes from recognition. Superresolution structured illumination microscopy (SR-SIM) revealed that the persistent phagosomal F-actin formed a cage-like structure that sterically hindered and functionally blocked the fusion of lysosomes. Blocking lysosome fusion was sufficient to inhibit phagosomal acidification and subsequent intracellular fungal killing by DCs. Retention of phagosomal F-actin by C. gattii also caused DC immunoparalysis. Disrupting the retained F-actin cage with cytochalasin D not only restored DC phagosomal maturation but also promoted DC costimulatory maturation and robust T cell activation and proliferation. Collectively, these results reveal a unique mechanism of DC immune evasion that enhances intracellular fungal pathogenicity and may explain suppressed cell-mediated immunity.IMPORTANCECryptococcus yeast species typically display characteristics of opportunistic pathogens, with the exception of C. gattii, which can cause life-threatening respiratory and disseminated brain infections in otherwise healthy people. The pathogenesis of C. gattii is not well understood, but an important characteristic is that C. gattii is capable of evading host cell-mediated immune defenses initiated by DCs. Here, we report that when virulent C. gattii becomes ingested by a DC, the intracellular compartment containing the fungi is covered by a persistent protein cage structure consisting of F-actin. This F-actin cage acts as a barrier to prevent interaction with other intracellular compartments, and as a result, the DC fails to kill the fungi and activate important cell-mediated immune responses. We propose that this unique immune evasion mechanism permits C. gattii to remain unchallenged within host cells, leading to persistent infection.
Keywords: Cryptococcus gattii; dendritic cells; immune evasion; immunoparalysis; phagosomal F-actin.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Kidd SE, Hagen F, Tscharke RL, Huynh M, Bartlett KH, Fyfe M, MacDougall L, Boekhout T, Kwon-Chung KJ, Meyer W. 2004. A rare genotype of Cryptococcus gattii caused the cryptococcosis outbreak on Vancouver Island (British Columbia, Canada). Proc Natl Acad Sci U S A 101:17258–17263. doi:10.1073/pnas.0402981101. - DOI - PMC - PubMed
-
- Harris JR, Lockhart SR, Sondermeyer G, Vugia DJ, Crist MB, D’Angelo MT, Sellers B, Franco-Paredes C, Makvandi M, Smelser C, Greene J, Stanek D, Signs K, Nett RJ, Chiller T, Park BJ. 2013. Cryptococcus gattii infections in multiple states outside the US Pacific Northwest. Emerg Infect Dis 19:1620–1626. doi:10.3201/eid1910.130441. - DOI - PMC - PubMed
-
- Leongson K, Cousineau-Cote V, Goupil M, Aumont F, Senechal S, Gaboury L, Jolicoeur P, Kronstad JW, de Repentigny L. 2013. Altered immune response differentially enhances susceptibility to Cryptococcus neoformans and Cryptococcus gattii infection in mice expressing the HIV-1 transgene. Infect Immun 81:1100–1113. doi:10.1128/IAI.01339-12. - DOI - PMC - PubMed
-
- Mody CH, Lipscomb MF, Street NE, Toews GB. 1990. Depletion of CD4+ (L3T4+) lymphocytes in vivo impairs murine host defense to Cryptococcus neoformans. J Immunol 144:1472–1477. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials