A Unified Model for the Function of YTHDF Proteins in Regulating m6A-Modified mRNA
- PMID: 32492408
- PMCID: PMC7508256
- DOI: 10.1016/j.cell.2020年05月01日2
A Unified Model for the Function of YTHDF Proteins in Regulating m6A-Modified mRNA
Abstract
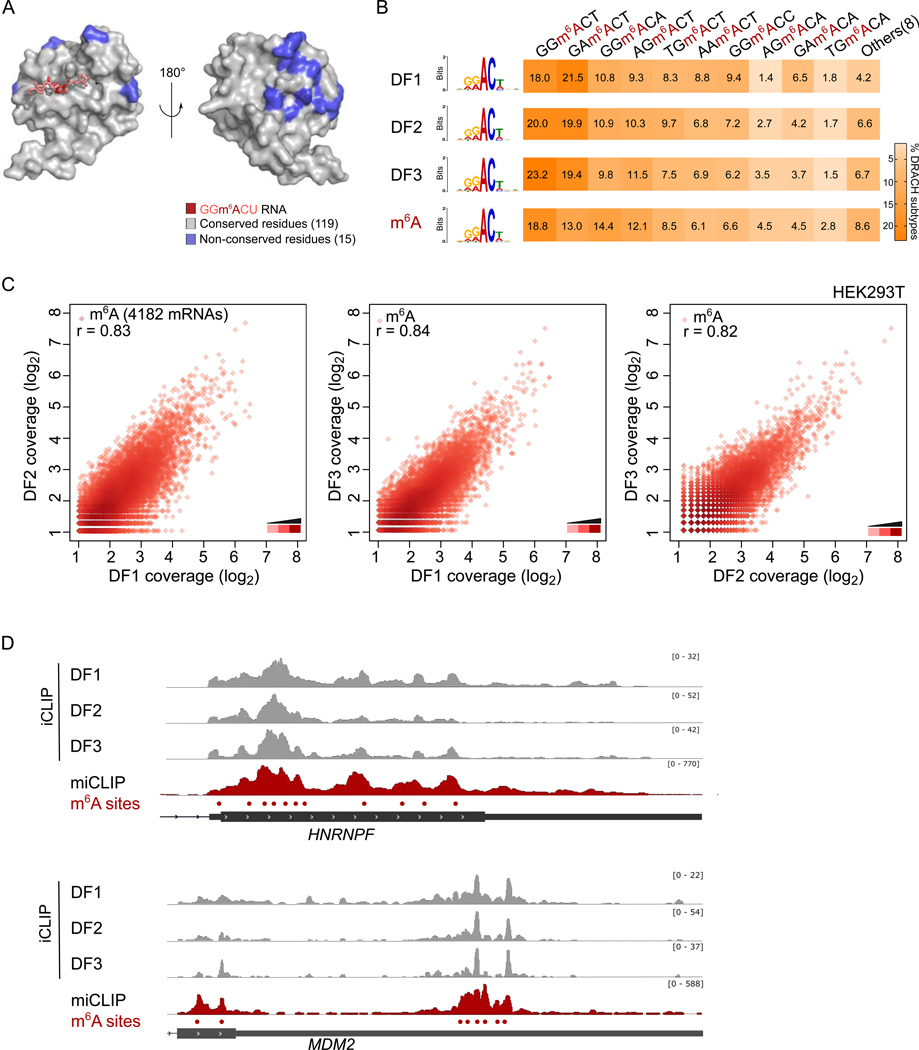
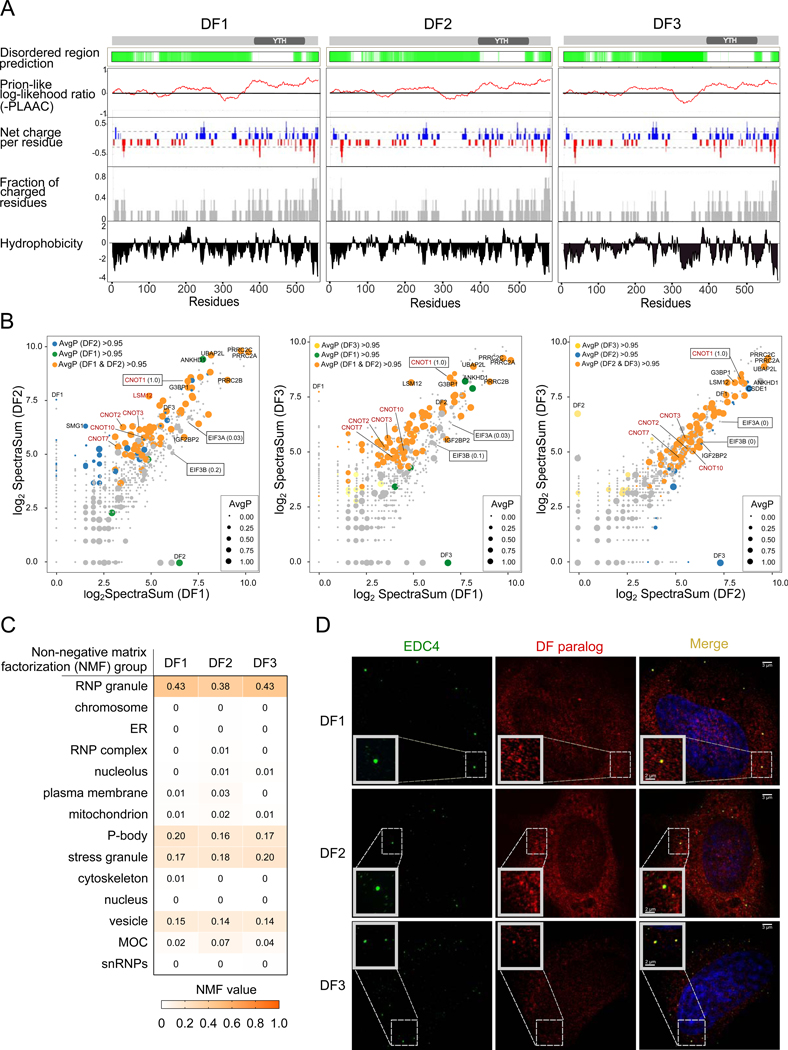
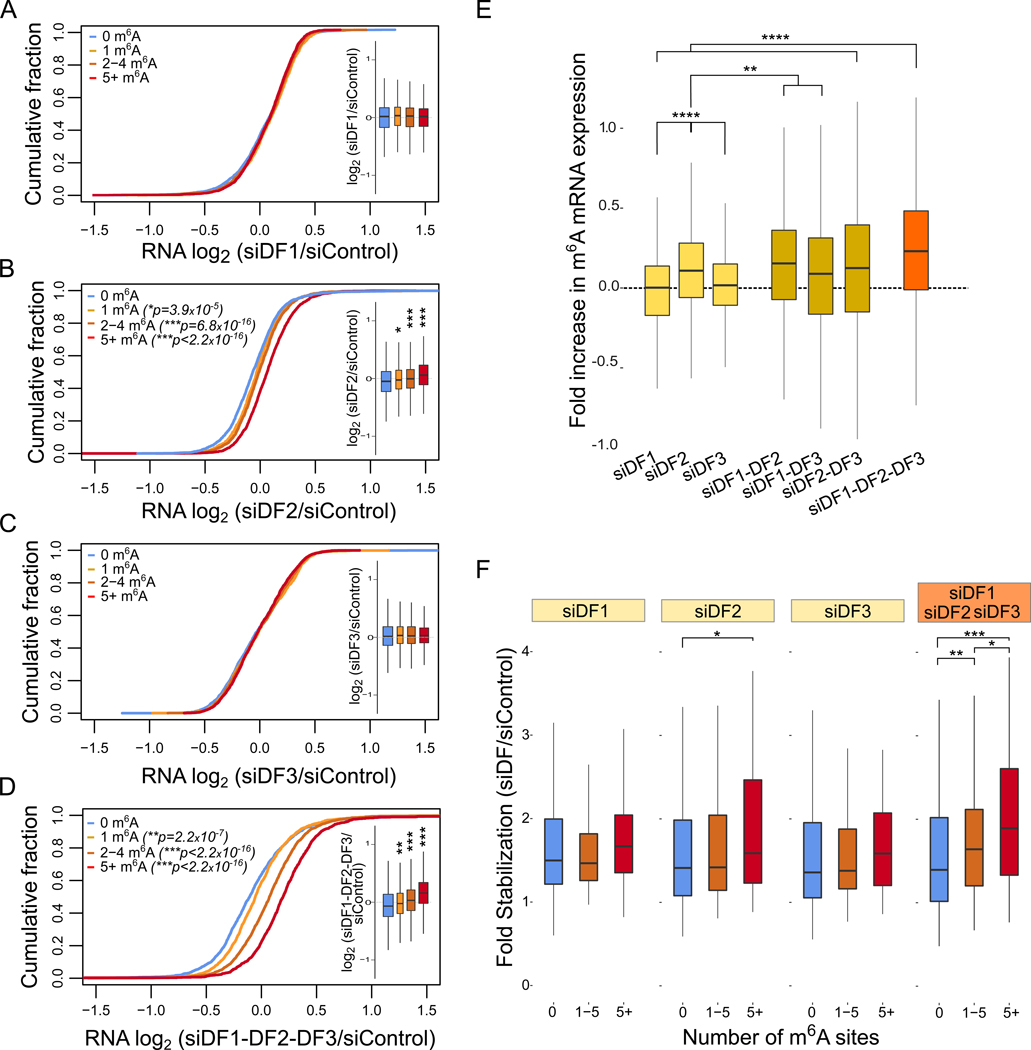
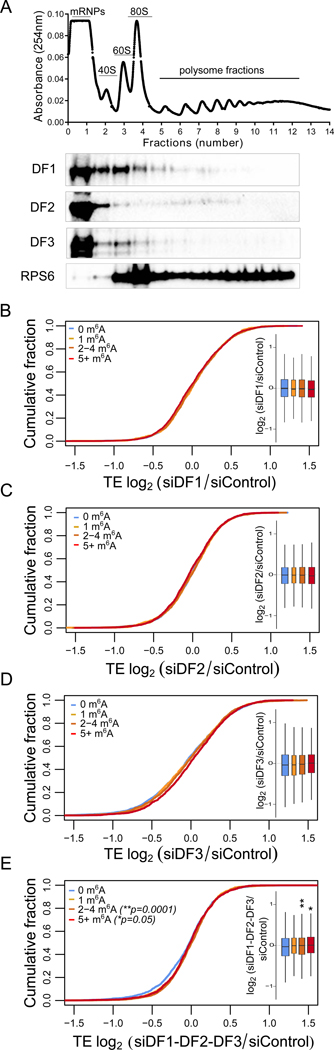
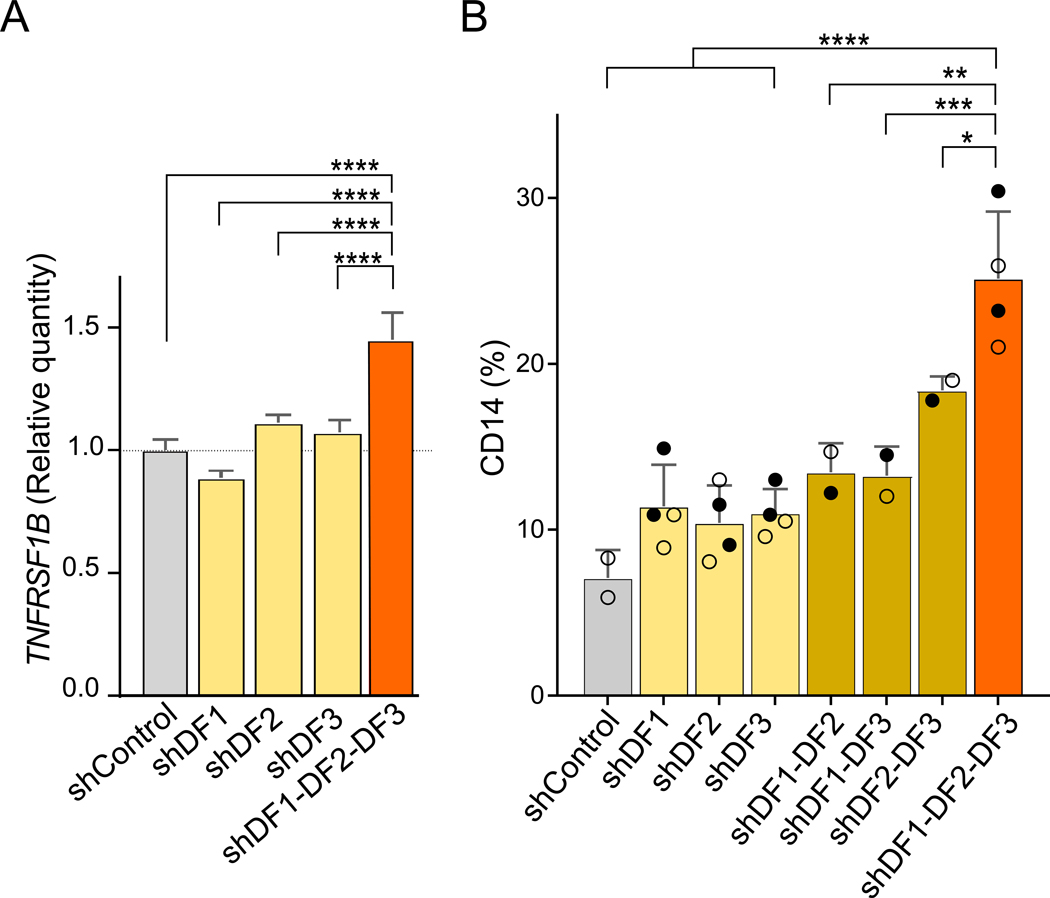
N6-methyladenosine (m6A) is the most abundant mRNA nucleotide modification and regulates critical aspects of cellular physiology and differentiation. m6A is thought to mediate its effects through a complex network of interactions between different m6A sites and three functionally distinct cytoplasmic YTHDF m6A-binding proteins (DF1, DF2, and DF3). In contrast to the prevailing model, we show that DF proteins bind the same m6A-modified mRNAs rather than different mRNAs. Furthermore, we find that DF proteins do not induce translation in HeLa cells. Instead, the DF paralogs act redundantly to mediate mRNA degradation and cellular differentiation. The ability of DF proteins to regulate stability and differentiation becomes evident only when all three DF paralogs are depleted simultaneously. Our study reveals a unified model of m6A function in which all m6A-modified mRNAs are subjected to the combined action of YTHDF proteins in proportion to the number of m6A sites.
Keywords: translation, mRNA stability, m(6)A, METTL3, YTHDF1, YTHDF2, YTHDF3, CLIP, RNA-binding.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests S.R.J. is scientific founder of, advisor to, and owns equity in Gotham Therapeutics.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous