5-fluorocytosine resistance is associated with hypermutation and alterations in capsule biosynthesis in Cryptococcus
- PMID: 31913284
- PMCID: PMC6949227
- DOI: 10.1038/s41467-019-13890-z
5-fluorocytosine resistance is associated with hypermutation and alterations in capsule biosynthesis in Cryptococcus
Abstract
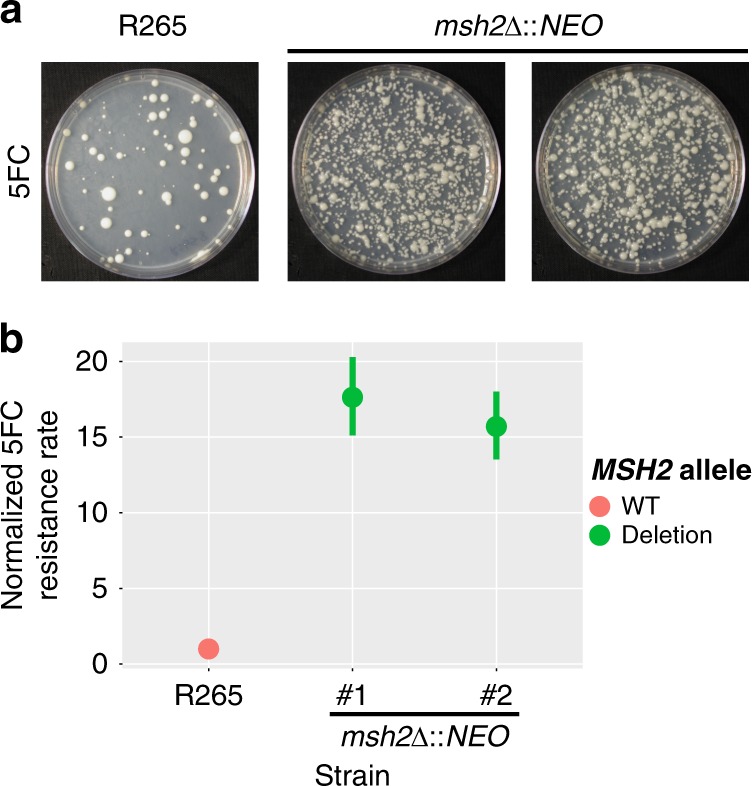
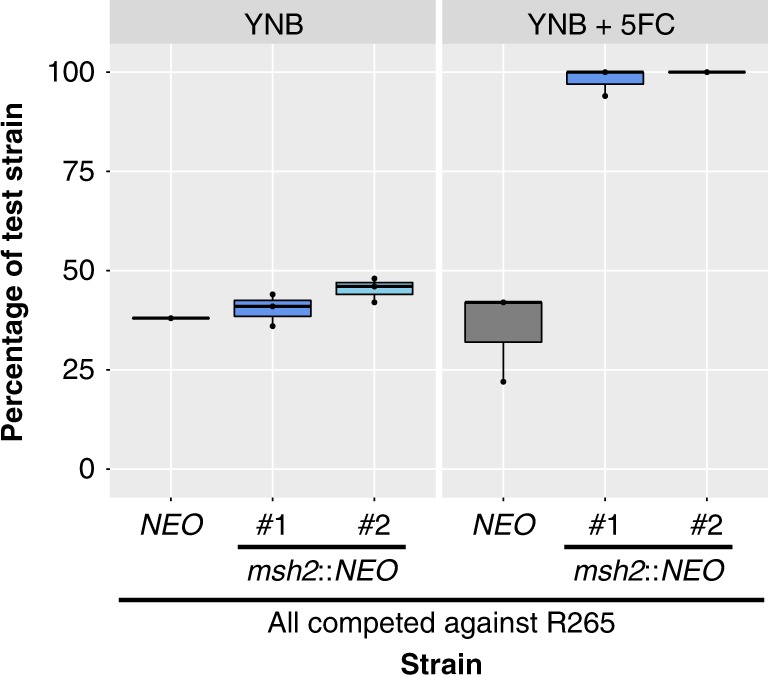
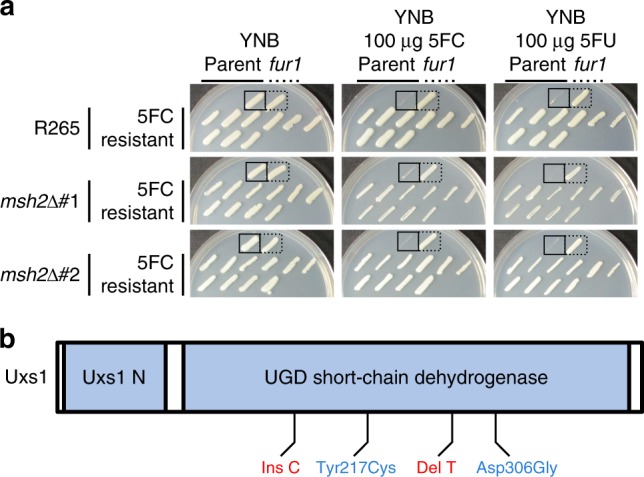
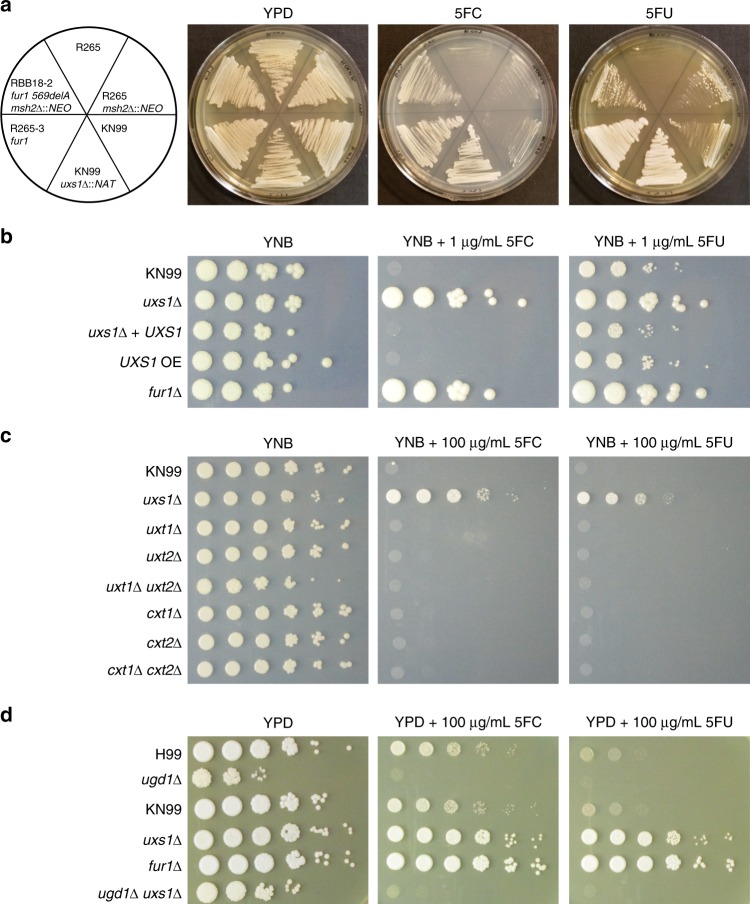
Patients infected with the fungal pathogen Cryptococcus are most effectively treated with a combination of 5-fluorocytosine (5FC) and amphotericin B. 5FC acts as a prodrug, which is converted into toxic 5-fluorouracil (5FU) upon uptake into fungal cells. However, the pathogen frequently develops resistance through unclear mechanisms. Here we show that resistance to 5FC in Cryptococcus deuterogattii is acquired more frequently in isolates with defects in DNA mismatch repair that confer an elevated mutation rate. We use whole genome sequencing of 16 independent isolates to identify mutations associated with 5FC resistance in vitro. We find mutations in known resistance genes (FUR1 and FCY2) and in a gene UXS1, previously shown to encode an enzyme that converts UDP-glucuronic acid to UDP-xylose for capsule biosynthesis, but not known to play a role in 5FC metabolism. Mutations in UXS1 lead to accumulation of UDP-glucuronic acid and alterations in nucleotide metabolism, which appear to suppress toxicity of both 5FC and its toxic derivative 5FU.
Conflict of interest statement
The authors declare no competing interests.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources