Competition and succession among coral endosymbionts
- PMID: 31788212
- PMCID: PMC6875658
- DOI: 10.1002/ece3.5749
Competition and succession among coral endosymbionts
Abstract
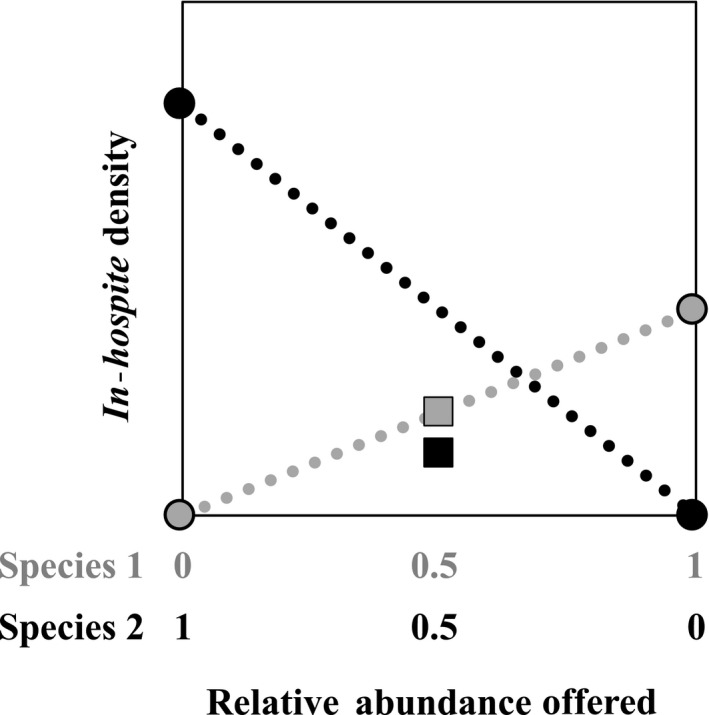
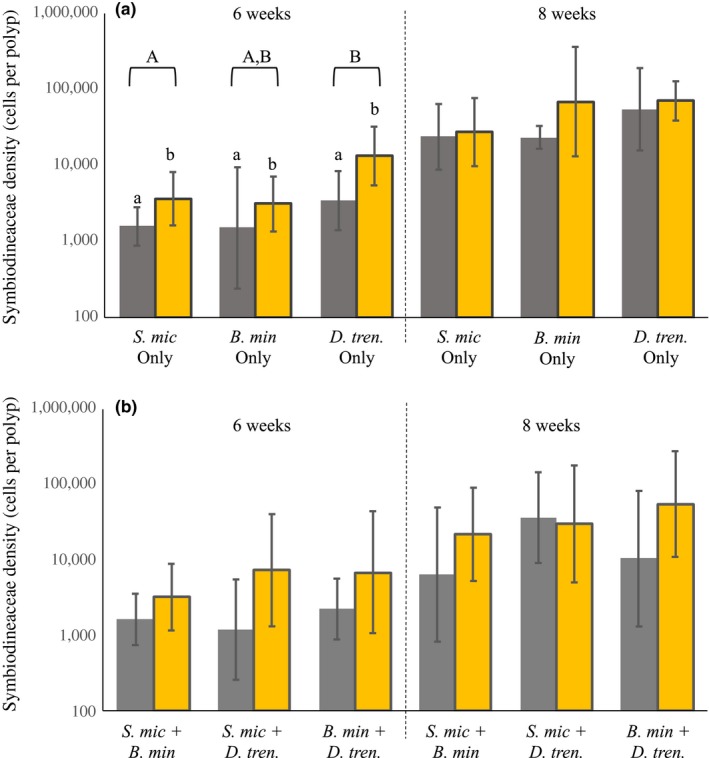
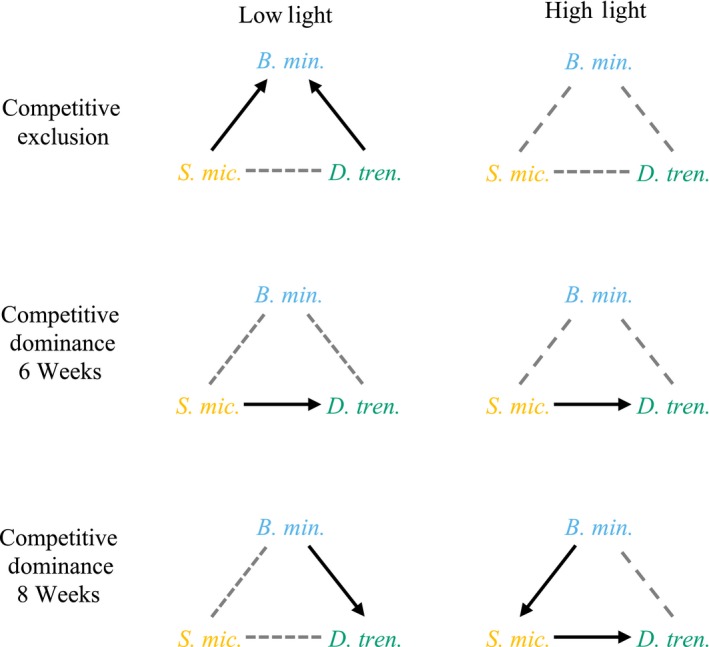
Host species often support a genetically diverse guild of symbionts, the identity and performance of which can determine holobiont fitness under particular environmental conditions. These symbiont communities are structured by a complex set of potential interactions, both positive and negative, between the host and symbionts and among symbionts. In reef-building corals, stable associations with specific symbiont species are common, and we hypothesize that this is partly due to ecological mechanisms, such as succession and competition, which drive patterns of symbiont winnowing in the initial colonization of new generations of coral recruits. We tested this hypothesis using the experimental framework of the de Wit replacement series and found that competitive interactions occurred among symbionts which were characterized by unique ecological strategies. Aposymbiotic octocoral recruits within high- and low-light environments were inoculated with one of three Symbiodiniaceae species as monocultures or with cross-paired mixtures, and we tracked symbiont uptake using quantitative genetic assays. Priority effects, in which early colonizers excluded competitive dominants, were evidenced under low light, but these early opportunistic species were later succeeded by competitive dominants. Under high light, a more consistent competitive hierarchy was established in which competitive dominants outgrew and limited the abundance of others. These findings provide insight into mechanisms of microbial community organization and symbiosis breakdown and recovery. Furthermore, transitions in competitive outcomes across spatial and temporal environmental variation may improve lifetime host fitness.
Keywords: Symbiodineaceae; competition; microbiome; succession; symbiosis.
© 2019 The Authors. Ecology and Evolution published by John Wiley & Sons Ltd.
Conflict of interest statement
All authors declare that there are no competing financial interests in relation to the work described.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Abrego, D. , Van Oppen, M. J. H. , & Willis, B. L. (2009). Onset of algal endosymbiont specificity varies among closely related species of Acropora corals during early ontogeny. Molecular Ecology, 18, 3532–3543. - PubMed
-
- Baker, A. (2003). Flexibility and specificity in coral‐algal symbiosis: Diversity, ecology, and biogeography of Symbiodinium . Annual Review of Ecology Evolution and Systematics, 34, 661–689.
LinkOut - more resources
Full Text Sources