A Novel Role of Fungal Type I Myosin in Regulating Membrane Properties and Its Association with d-Amino Acid Utilization in Cryptococcus gattii
- PMID: 31455652
- PMCID: PMC6712397
- DOI: 10.1128/mBio.01867-19
A Novel Role of Fungal Type I Myosin in Regulating Membrane Properties and Its Association with d-Amino Acid Utilization in Cryptococcus gattii
Abstract
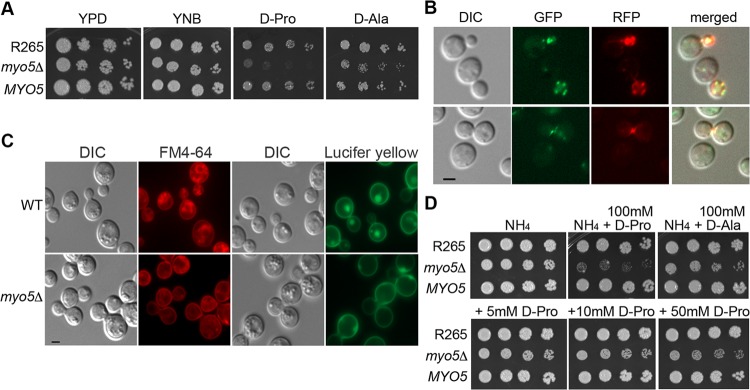
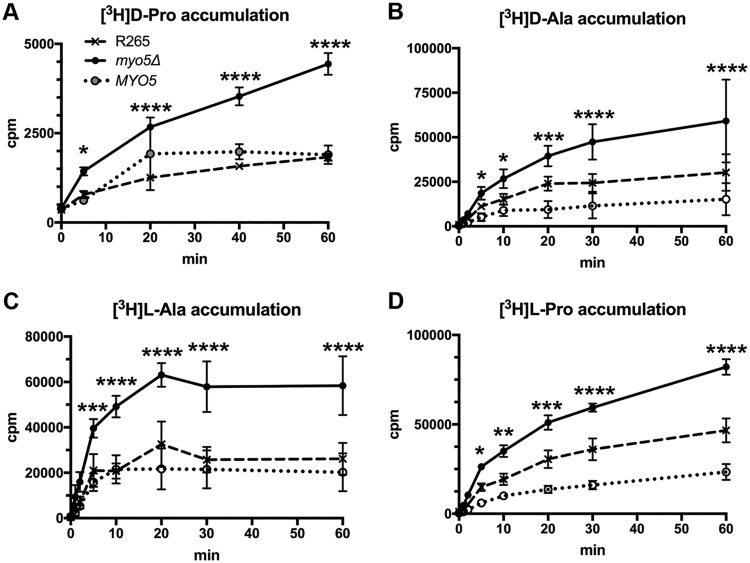
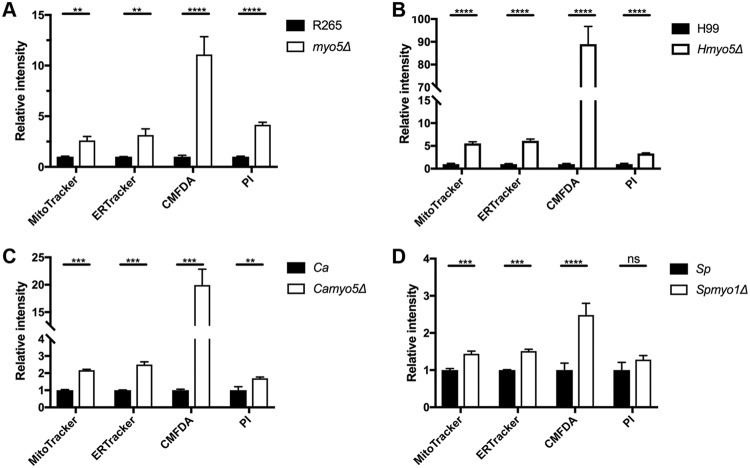
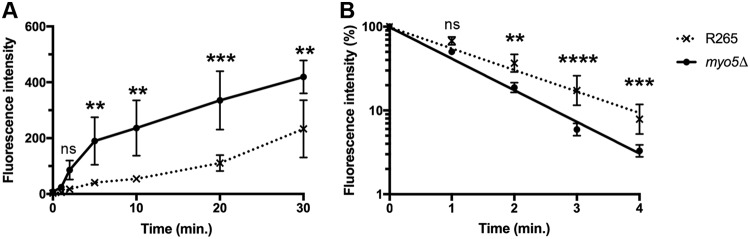
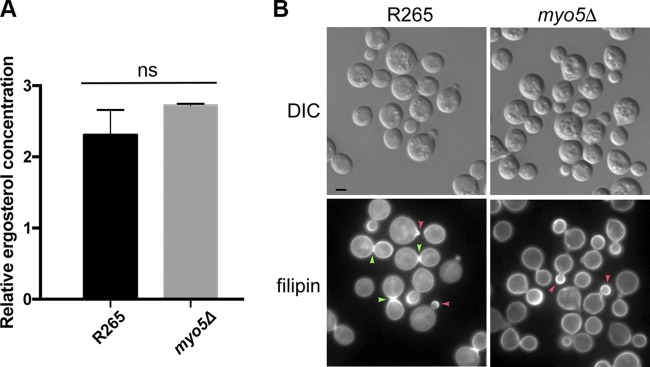
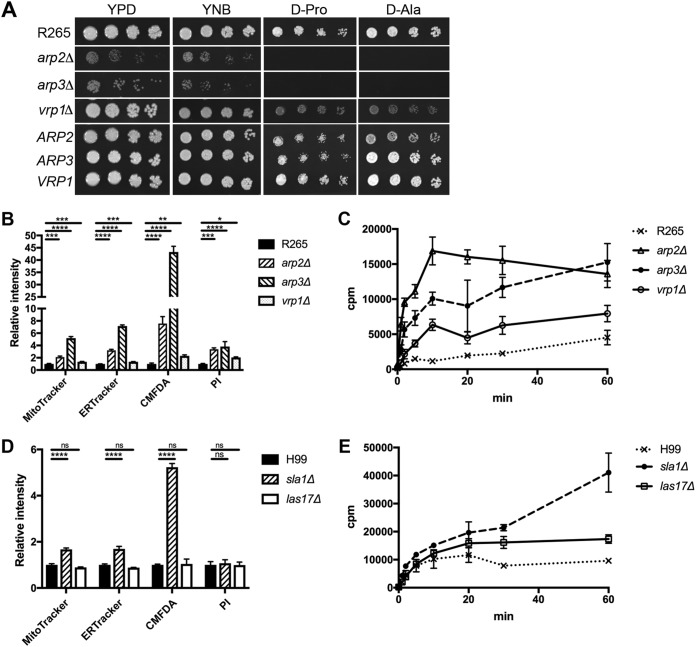
We found a novel role of Myo5, a type I myosin (myosin-I), and its fortuitous association with d-amino acid utilization in Cryptococcus gattii Myo5 colocalized with actin cortical patches and was required for endocytosis. Interestingly, the myo5Δ mutant accumulated high levels of d-proline and d-alanine which caused toxicity in C. gattii cells. The myo5Δ mutant also accumulated a large set of substrates, such as membrane-permeant as well as non-membrane-permeant dyes, l-proline, l-alanine, and flucytosine intracellularly. Furthermore, the efflux rate of fluorescein was significantly increased in the myo5Δ mutant. Importantly, the endocytic defect of the myo5Δ mutant did not affect the localization of the proline permease and flucytosine transporter. These data indicate that the substrate accumulation phenotype is not solely due to a defect in endocytosis, but the membrane properties may have been altered in the myo5Δ mutant. Consistent with this, the sterol staining pattern of the myo5Δ mutant was different from that of the wild type, and the mutant was hypersensitive to amphotericin B. It appears that the changes in sterol distribution may have caused altered membrane permeability in the myo5Δ mutant, allowing increased accumulation of substrate. Moreover, myosin-I mutants generated in several other yeast species displayed a similar substrate accumulation phenotype. Thus, fungal type I myosin appears to play an important role in regulating membrane permeability. Although the substrate accumulation phenotype was detected in strains with mutations in the genes involved in actin nucleation, the phenotype was not shared in all endocytic mutants, indicating a complicated relationship between substrate accumulation and endocytosis.IMPORTANCECryptococcus gattii, one of the etiological agents of cryptococcosis, can be distinguished from its sister species Cryptococcus neoformans by growth on d-amino acids. C. gattiiMYO5 affected the growth of C. gattii on d-amino acids. The myo5Δ cells accumulated high levels of various substrates from outside the cells, and excessively accumulated d-amino acids appeared to have caused toxicity in the myo5Δ cells. We provide evidence on the alteration of membrane properties in the myo5Δ mutants. Additionally, alteration in the myo5Δ membrane permeability causing higher substrate accumulation is associated with the changes in the sterol distribution. Furthermore, myosin-I in three other yeasts also manifested a similar role in substrate accumulation. Thus, while fungal myosin-I may function as a classical myosin-I, it has hitherto unknown additional roles in regulating membrane permeability. Since deletion of fungal myosin-I causes significantly elevated susceptibility to multiple antifungal drugs, it could serve as an effective target for augmentation of fungal therapy.
Keywords: Cryptococcus; actin patches; endocytosis; membrane permeability; myosin-I.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Coluccio LM. 2008. Myosin I, p 95–124. In Coluccio LM. (ed), Myosins: a superfamily of molecular motors. Springer, Dordrecht, The Netherlands. doi:10.1007/978-1-4020-6519-4. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources