Molecular evidence of sequential evolution of DDT- and pyrethroid-resistant sodium channel in Aedes aegypti
- PMID: 31158225
- PMCID: PMC6564045
- DOI: 10.1371/journal.pntd.0007432
Molecular evidence of sequential evolution of DDT- and pyrethroid-resistant sodium channel in Aedes aegypti
Abstract
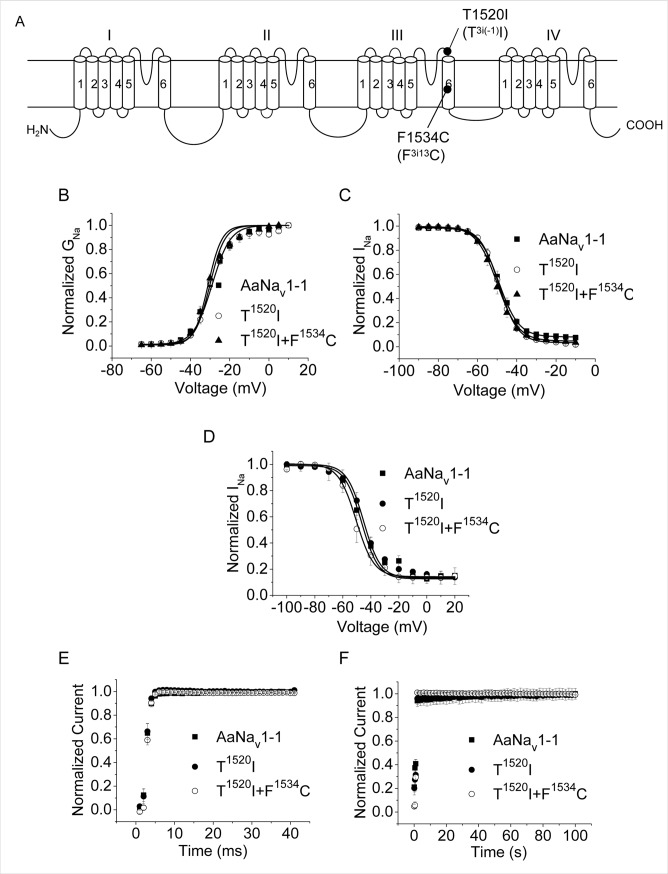
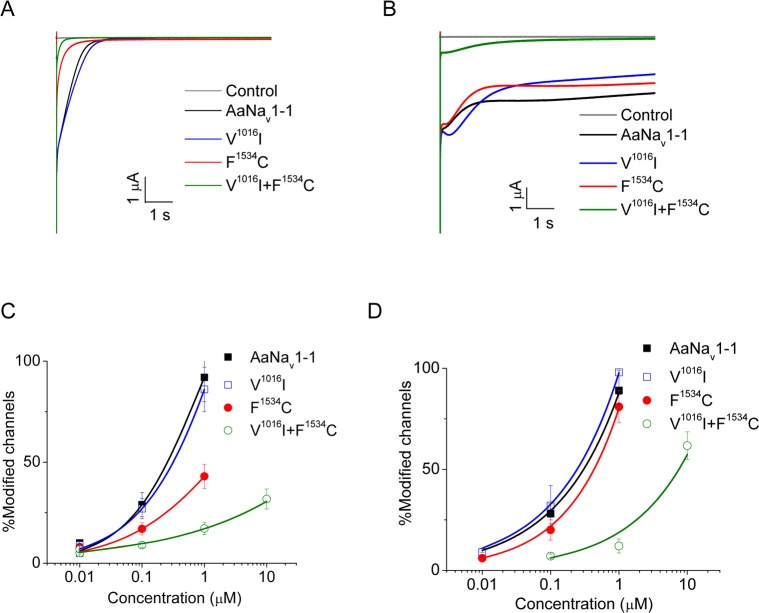
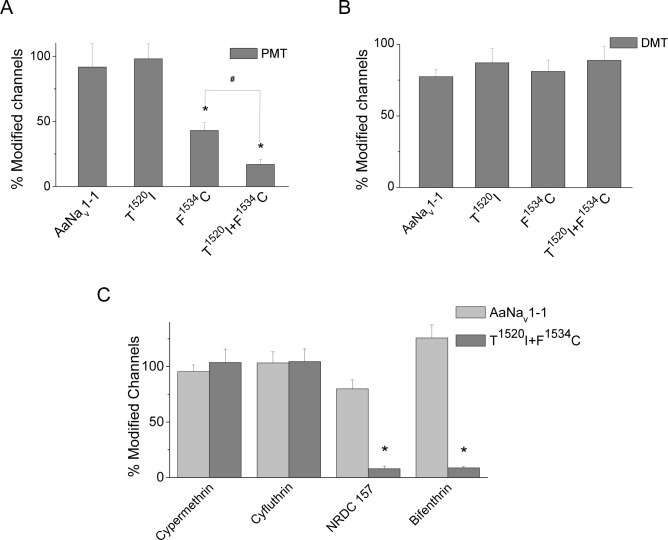
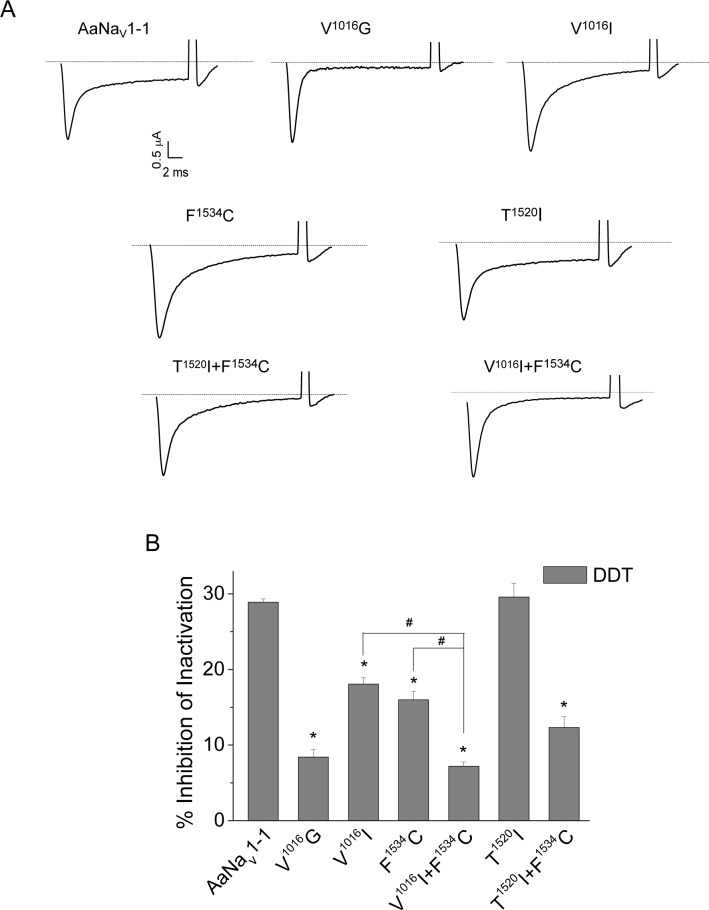
Background: Multiple mutations in the voltage-gated sodium channel have been associated with knockdown resistance (kdr) to DDT and pyrethroid insecticides in a major human disease vector Aedes aegypti. One mutation, V1016G, confers sodium channel resistance to pyrethroids, but a different substitution in the same position V1016I alone had no effect. In pyrethroid-resistant Ae. aegypti populations, V1016I is often linked to another mutation, F1534C, which confers sodium channel resistance only to Type I pyrethroids including permethrin (PMT), but not to Type II pyrethroids including deltamethrin (DMT). Mosquitoes carrying both V1016G and F1534C exhibited a greater level of pyrethroid resistance than those carrying F1534C alone. More recently, a new mutation T1520I co-existing with F1534C was detected in India. However, whether V1016I or T1520I enhances pyrethroid resistance of sodium channels carrying F1534C remains unknown.
Methodology/principal findings: V1016I, V1016G, T1520I and F1534C substitutions were introduced alone and in various combinations into AaNav1-1, a sodium channel from Aedes aegypti. The mutant channels were then expressed in Xenopus oocytes and examined for channel properties and sensitivity to pyrethroids using the two-electrode voltage clamping technique. The results showed that V1016I or T1520I alone did not alter the AaNav1-1 sensitivity to PMT or DMT. However, the double mutant T1520I+F1534C was more resistant to PMT than F1534C, but remained sensitive to DMT. In contrast, the double mutant V1016I+F1534C was resistant to DMT and more resistant to PMT than F1534C. Furthermore, V1016I/G and F1534C channels, but not T1520I, were resistant to dichlorodiphenyltrichloroethane (DDT). Cryo-EM structures of sodium channels suggest that T1520I allosterically deforms geometry of the pyrethroid receptor site PyR1 in AaNav1-1. The small deformation does not affect binding of DDT, PMT or DMT, but in combination with F1534C it increases the channel resistance to PMT and DDT.
Conclusions/significance: Our data corroborated the previously proposed sequential selection of kdr mutations in Ae. aegypti. We proposed that mutation F1534C first emerged in response to DDT/pyrethroids providing a platform for subsequent selection of mutations V1016I and T1520I that confer greater and broader spectrum of pyrethroid resistance.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Elliott M. Synthetic pyrethroids. Elliott M, editor. Washington, D.C.: American Chemical Society; 1977. 1–28 p.
-
- Goldin AL. Mechanisms of sodium channel inactivation. Curr Opin Neurobiol. 2003;13(3):284–90. Epub 2003年07月10日. S0959438803000655 [pii]. . - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous