Sexual reproduction in a natural Trypanosoma cruzi population
- PMID: 31107905
- PMCID: PMC6544315
- DOI: 10.1371/journal.pntd.0007392
Sexual reproduction in a natural Trypanosoma cruzi population
Abstract
Background: Sexual reproduction provides an evolutionary advantageous mechanism that combines favorable mutations that have arisen in separate lineages into the same individual. This advantage is especially pronounced in microparasites as allelic reassortment among individuals caused by sexual reproduction promotes allelic diversity at immune evasion genes within individuals which is often essential to evade host immune systems. Despite these advantages, many eukaryotic microparasites exhibit highly-clonal population structures suggesting that genetic exchange through sexual reproduction is rare. Evidence supporting clonality is particularly convincing in the causative agent of Chagas disease, Trypanosoma cruzi, despite equally convincing evidence of the capacity to engage in sexual reproduction.
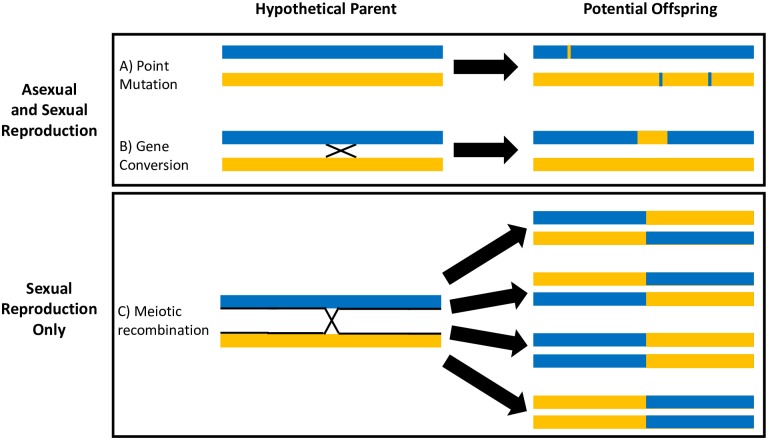
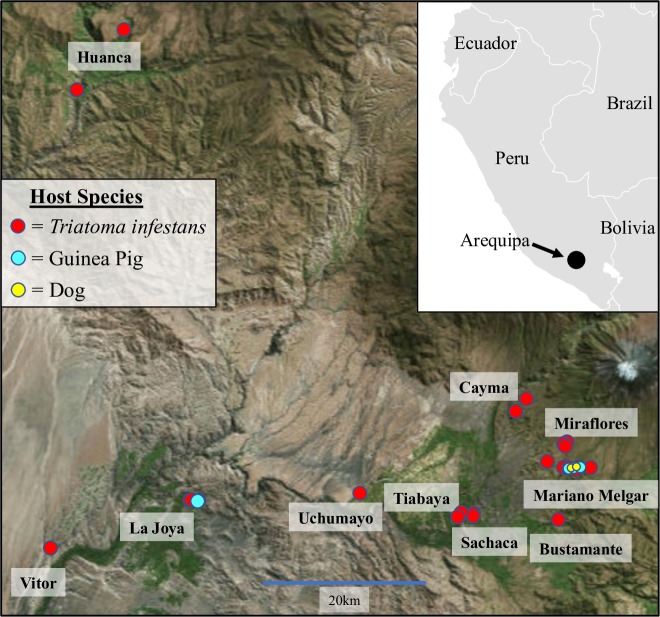
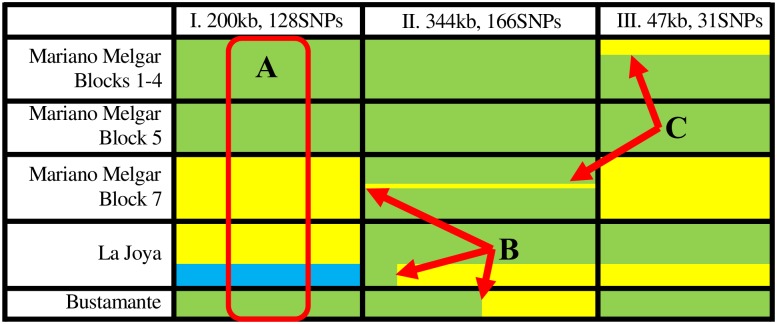
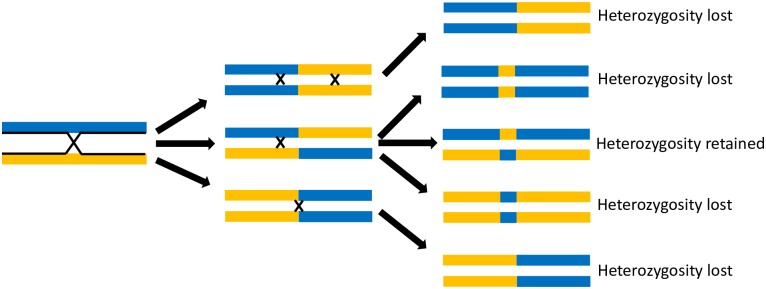
Methodology/ principle findings: In the present study, we investigated two hypotheses that can reconcile the apparent contradiction between the observed clonal population structure and the capacity to engage in sexual reproduction by analyzing the genome sequences of 123 T. cruzi isolates from a natural population in Arequipa, Peru. The distribution of polymorphic markers within and among isolates provides clear evidence of the occurrence of sexual reproduction. Large genetic segments are rearranged among chromosomes due to crossing over during meiosis leading to a decay in the genetic linkage among polymorphic markers compared to the expectations from a purely asexually-reproducing population. Nevertheless, the population structure appears clonal due to a high level of inbreeding during sexual reproduction which increases homozygosity, and thus reduces diversity, within each inbreeding lineage.
Conclusions/ significance: These results effectively reconcile the apparent contradiction by demonstrating that the clonal population structure is derived not from infrequent sex in natural populations but from high levels of inbreeding. We discuss epidemiological consequences of this reproductive strategy on genome evolution, population structure, and phenotypic diversity of this medically important parasite.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Mihok S, Otieno LH, Darji N. Population genetics of Trypanosoma brucei and the epidemiology of human sleeping sickness in the Lambwe Valley, Kenya. Parasitology: Cambridge University Press; 1990;100: 219–233. - PubMed
-
- Tibayrenc M, Kjellberg F, Ayala FJ. A clonal theory of parasitic protozoa: the population structures of Entamoeba, Giardia, Leishmania, Naegleria, Plasmodium, Trichomonas, and Trypanosoma and their medical and taxonomical consequences. Proceedings of the National Academy of Sciences of the United States of America. 1990;87: 2414–2418. 10.1073/pnas.87.7.2414 - DOI - PMC - PubMed
-
- Pujol C, Reynes J, Renaud F, Raymond M, Tibayrenc M, Ayala FJ, et al. The yeast Candida albicans has a clonal mode of reproduction in a population of infected human immunodeficiency virus-positive patients. Proceedings of the National Academy of Sciences. National Acad Sciences; 1993;90: 9456–9459. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical