A Novel Protocol for the Isolation of Fungal Extracellular Vesicles Reveals the Participation of a Putative Scramblase in Polysaccharide Export and Capsule Construction in Cryptococcus gattii
- PMID: 30894430
- PMCID: PMC6429041
- DOI: 10.1128/mSphere.00080-19
A Novel Protocol for the Isolation of Fungal Extracellular Vesicles Reveals the Participation of a Putative Scramblase in Polysaccharide Export and Capsule Construction in Cryptococcus gattii
Abstract
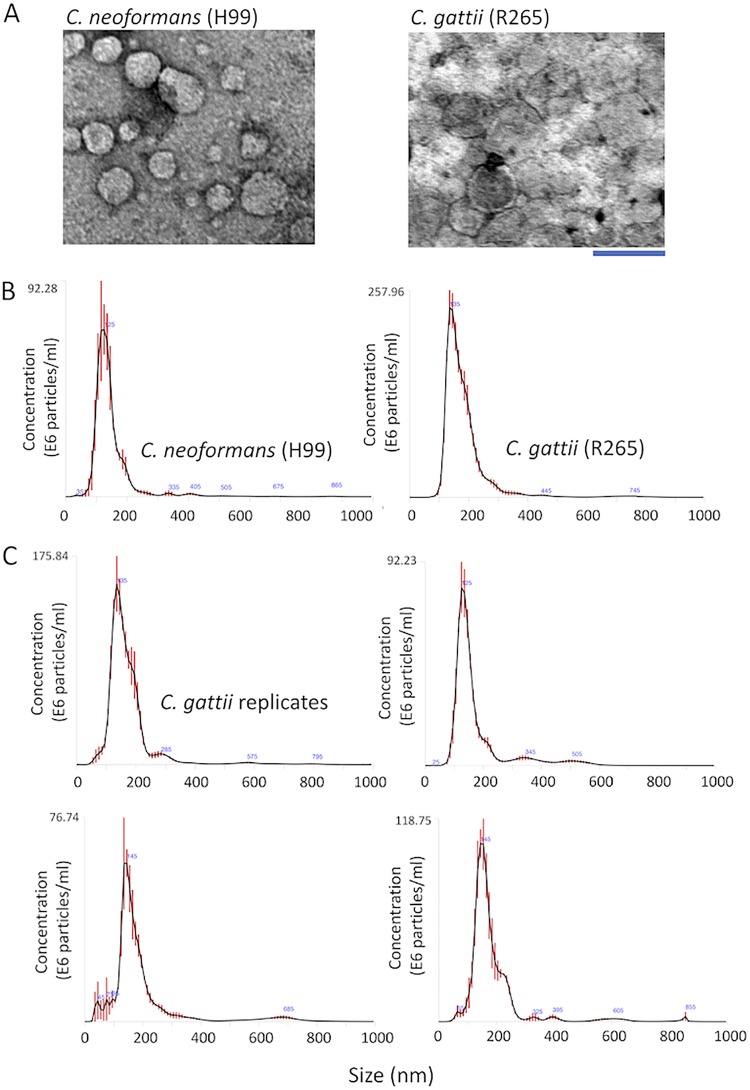
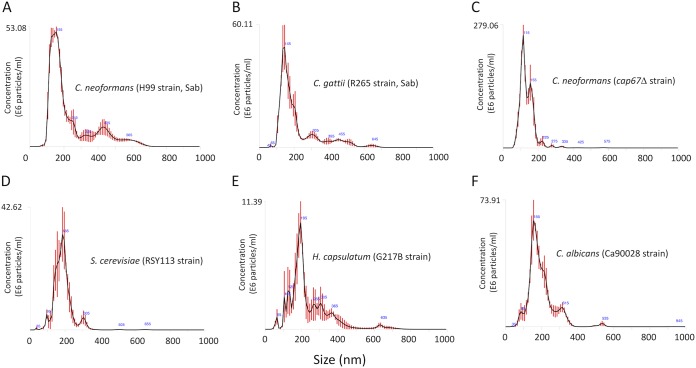
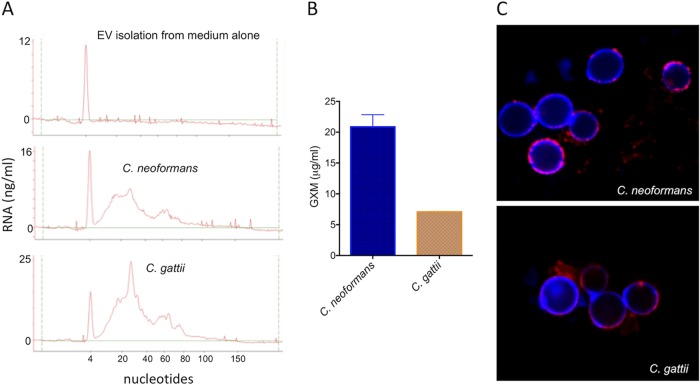
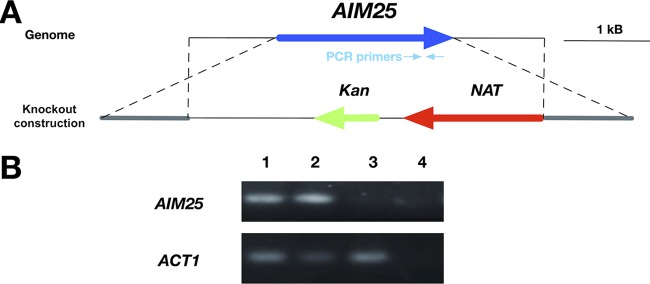
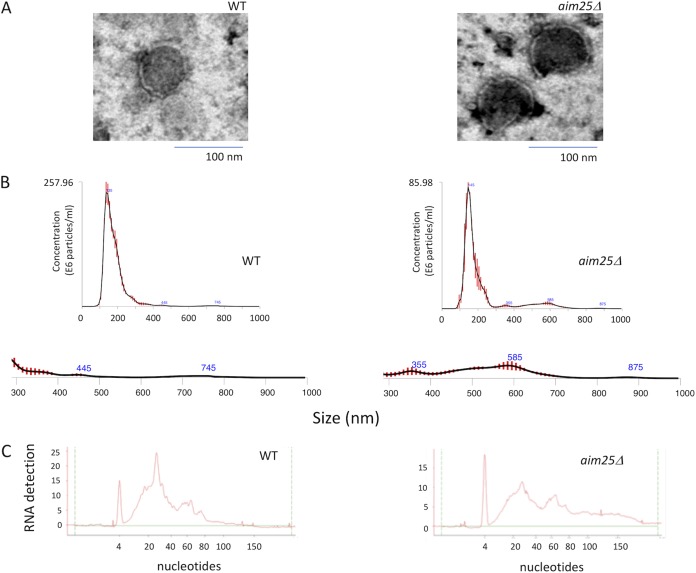
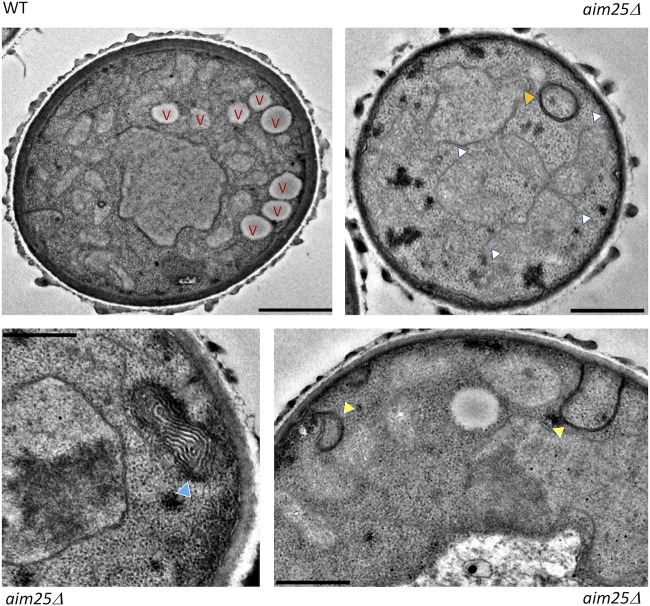
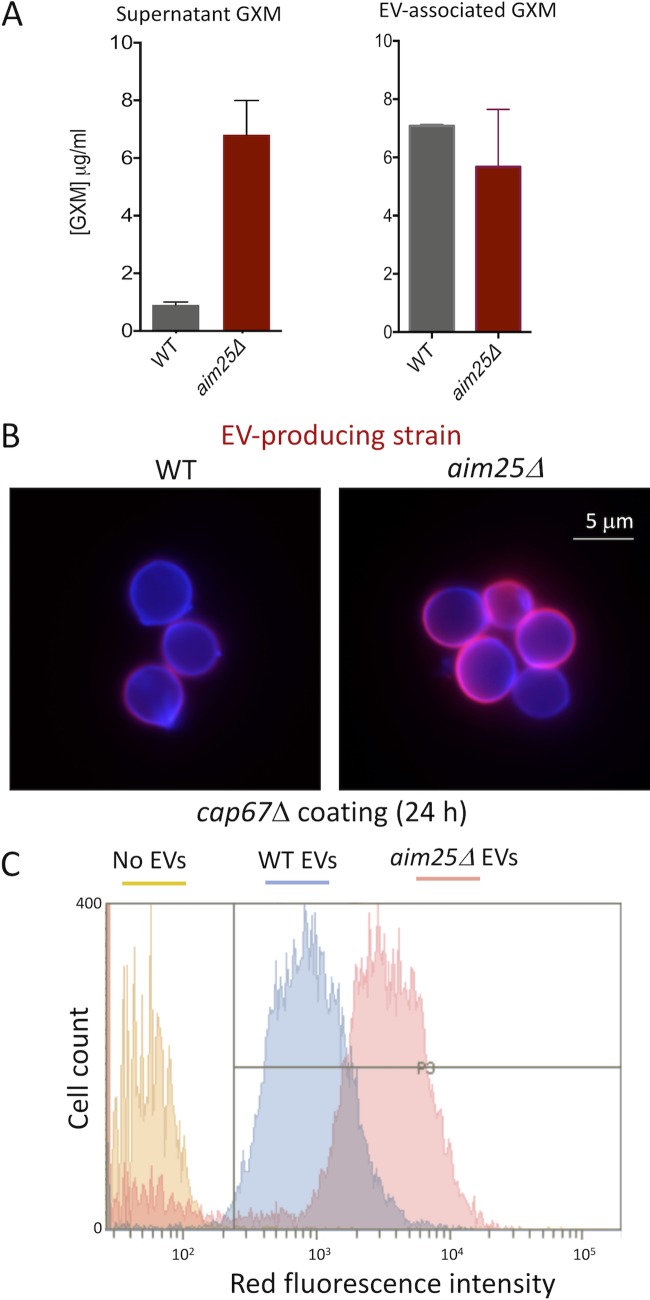
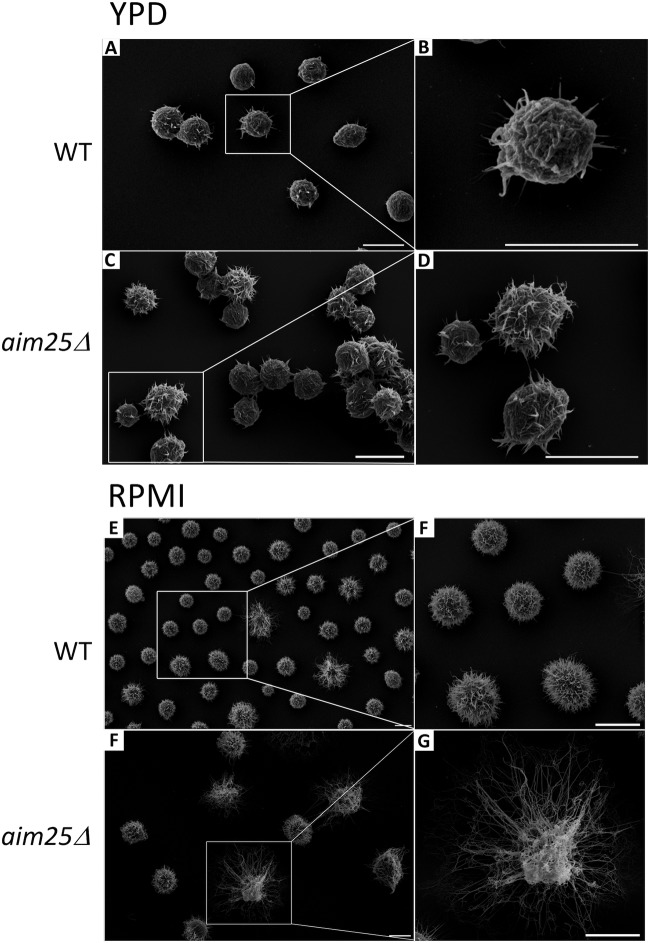
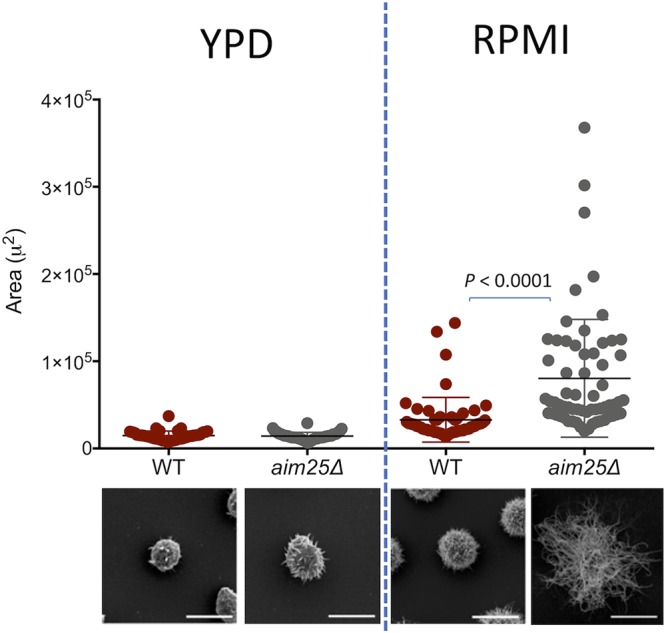
Regular protocols for the isolation of fungal extracellular vesicles (EVs) are time-consuming, hard to reproduce, and produce low yields. In an attempt to improve the protocols used for EV isolation, we explored a model of vesicle production after growth of Cryptococcus gattii and Cryptococcus neoformans on solid media. Nanoparticle tracking analysis in combination with transmission electron microscopy revealed that C. gattii and C. neoformans produced EVs in solid media. The properties of cryptococcal vesicles varied according to the culture medium used and the EV-producing species. EV detection was reproduced with an acapsular mutant of C. neoformans, as well as with isolates of Candida albicans, Histoplasma capsulatum, and Saccharomyces cerevisiae Cryptococcal EVs produced in solid media were biologically active and contained regular vesicular components, including the major polysaccharide glucuronoxylomannan (GXM) and RNA. Since the protocol had higher yields and was much faster than the regular methods used for the isolation of fungal EVs, we asked if it would be applicable to address fundamental questions related to cryptococcal secretion. On the basis that polysaccharide export in Cryptococcus requires highly organized membrane traffic culminating with EV release, we analyzed the participation of a putative scramblase (Aim25; CNBG_3981) in EV-mediated GXM export and capsule formation in C. gattii EVs from a C. gattiiaim25Δ strain differed from those obtained from wild-type (WT) cells in physical-chemical properties and cargo. In a model of surface coating of an acapsular cryptococcal strain with vesicular GXM, EVs obtained from the aim25Δ mutant were more efficiently used as a source of capsular polysaccharides. Lack of the Aim25 scramblase resulted in disorganized membranes and increased capsular dimensions. These results associate the description of a novel protocol for the isolation of fungal EVs with the identification of a previously unknown regulator of polysaccharide release.IMPORTANCE Extracellular vesicles (EVs) are fundamental components of the physiology of cells from all kingdoms. In pathogenic fungi, they participate in important mechanisms of transfer of antifungal resistance and virulence, as well as in immune stimulation and prion transmission. However, studies on the functions of fungal EVs are still limited by the lack of efficient methods for isolation of these compartments. In this study, we developed an alternative protocol for isolation of fungal EVs and demonstrated an application of this new methodology in the study of the physiology of the fungal pathogen Cryptococcus gattii Our results describe a fast and reliable method for the study of fungal EVs and reveal the participation of scramblase, a phospholipid-translocating enzyme, in secretory processes of C. gattii.
Keywords: Cryptococcus; extracellular vesicles; fungi; scramblase; secretion.
Copyright © 2019 Reis et al.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Rodrigues ML, Nimrichter L, Oliveira DL, Frases S, Miranda K, Zaragoza O, Alvarez M, Nakouzi A, Feldmesser M, Casadevall A. 2007. Vesicular polysaccharide export in Cryptococcus neoformans is a eukaryotic solution to the problem of fungal trans-cell wall transport. Eukaryot Cell 6:48–59. doi:10.1128/EC.00318-06. - DOI - PMC - PubMed
-
- Zarnowski R, Sanchez H, Covelli AS, Dominguez E, Jaromin A, Bernhardt J, Mitchell KF, Heiss C, Azadi P, Mitchell A, Andes DR. 2018. Candida albicans biofilm-induced vesicles confer drug resistance through matrix biogenesis. PLoS Biol 16:e2006872. doi:10.1371/journal.pbio.2006872. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources