Genomic Analysis of Plasmodium vivax in Southern Ethiopia Reveals Selective Pressures in Multiple Parasite Mechanisms
- PMID: 30668735
- PMCID: PMC6804337
- DOI: 10.1093/infdis/jiz016
Genomic Analysis of Plasmodium vivax in Southern Ethiopia Reveals Selective Pressures in Multiple Parasite Mechanisms
Abstract
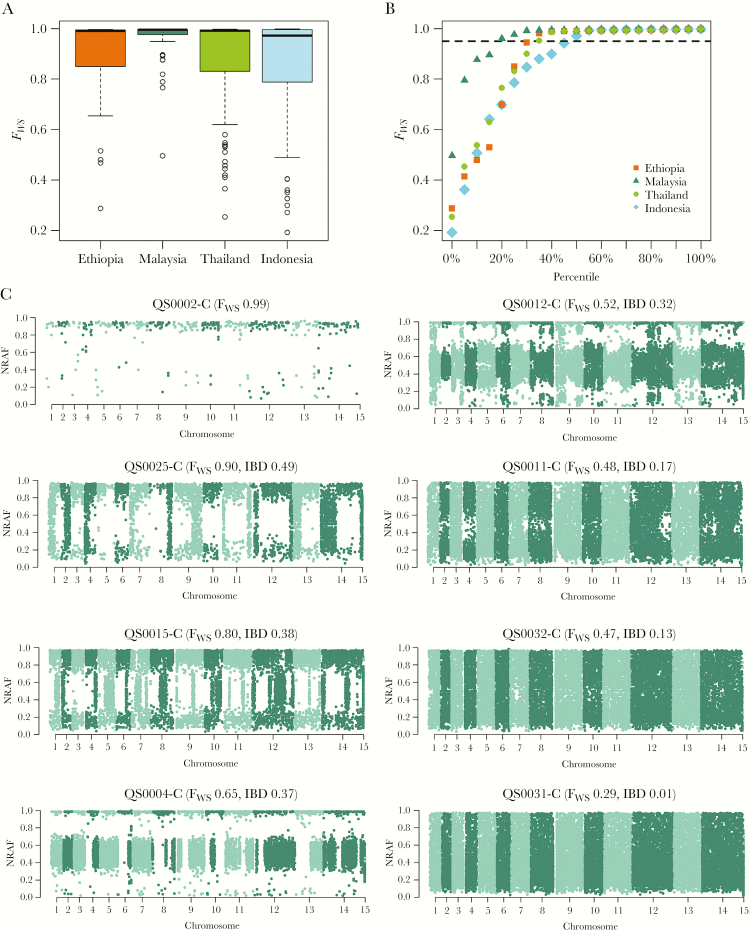
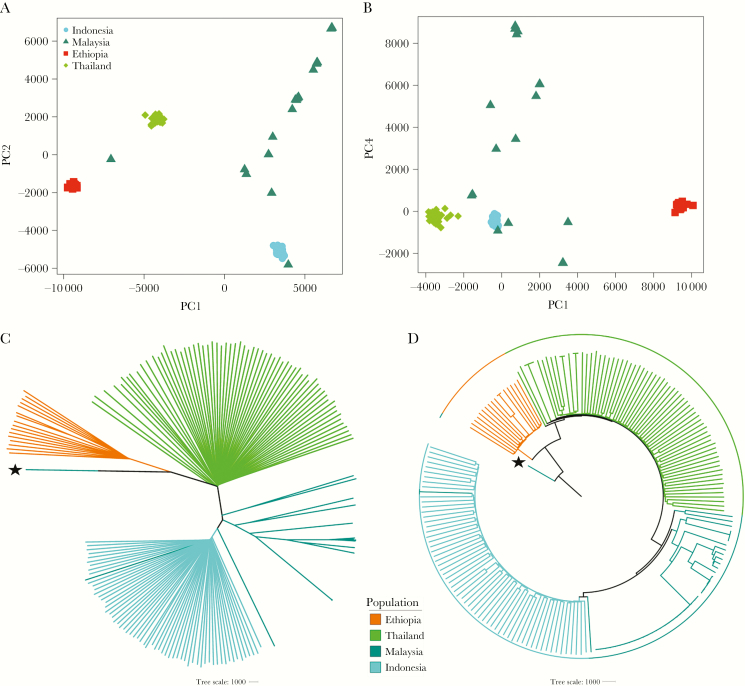
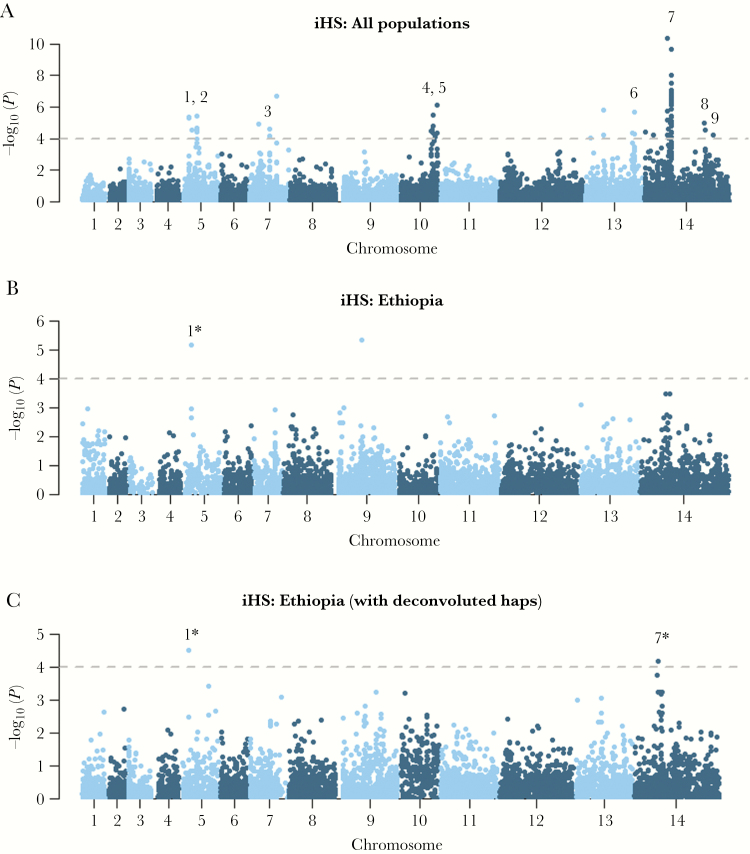
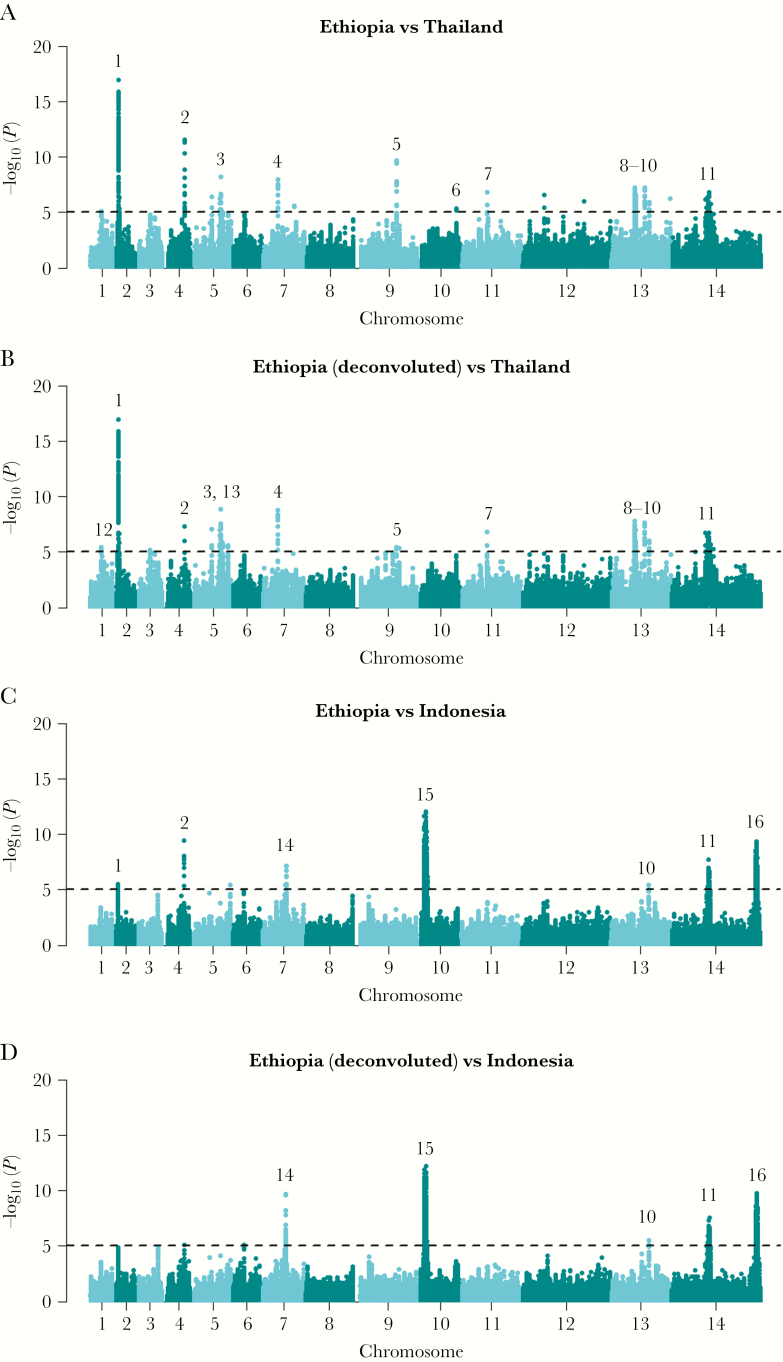
The Horn of Africa harbors the largest reservoir of Plasmodium vivax in the continent. Most of sub-Saharan Africa has remained relatively vivax-free due to a high prevalence of the human Duffy-negative trait, but the emergence of strains able to invade Duffy-negative reticulocytes poses a major public health threat. We undertook the first population genomic investigation of P. vivax from the region, comparing the genomes of 24 Ethiopian isolates against data from Southeast Asia to identify important local adaptions. The prevalence of the Duffy binding protein amplification in Ethiopia was 79%, potentially reflecting adaptation to Duffy negativity. There was also evidence of selection in a region upstream of the chloroquine resistance transporter, a putative chloroquine-resistance determinant. Strong signals of selection were observed in genes involved in immune evasion and regulation of gene expression, highlighting the need for a multifaceted intervention approach to combat P. vivax in the region.
Keywords: Plasmodium; Duffy; Ethiopia; genomics; malaria; vivax.
© The Author(s) 2019. Published by Oxford University Press for the Infectious Diseases Society of America.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Comment in
-
A Solid Beginning to Understanding Plasmodium vivax in Africa.Sibley CH. Sibley CH. J Infect Dis. 2019 Oct 22;220(11):1716-1718. doi: 10.1093/infdis/jiz019. J Infect Dis. 2019. PMID: 30668773 No abstract available.
References
-
- Sattabongkot J, Tsuboi T, Zollner GE, Sirichaisinthop J, Cui L. Plasmodium vivax transmission: chances for control? Trends Parasitol 2004; 20:192–8. - PubMed