Activity of acetyltransferase toxins involved in Salmonella persister formation during macrophage infection
- PMID: 29777131
- PMCID: PMC5959882
- DOI: 10.1038/s41467-018-04472-6
Activity of acetyltransferase toxins involved in Salmonella persister formation during macrophage infection
Abstract
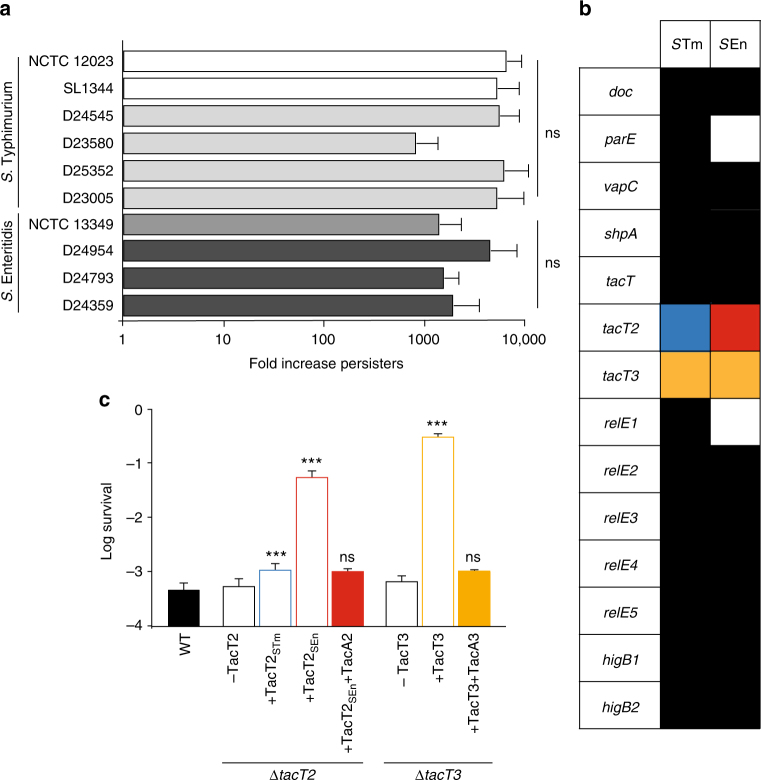
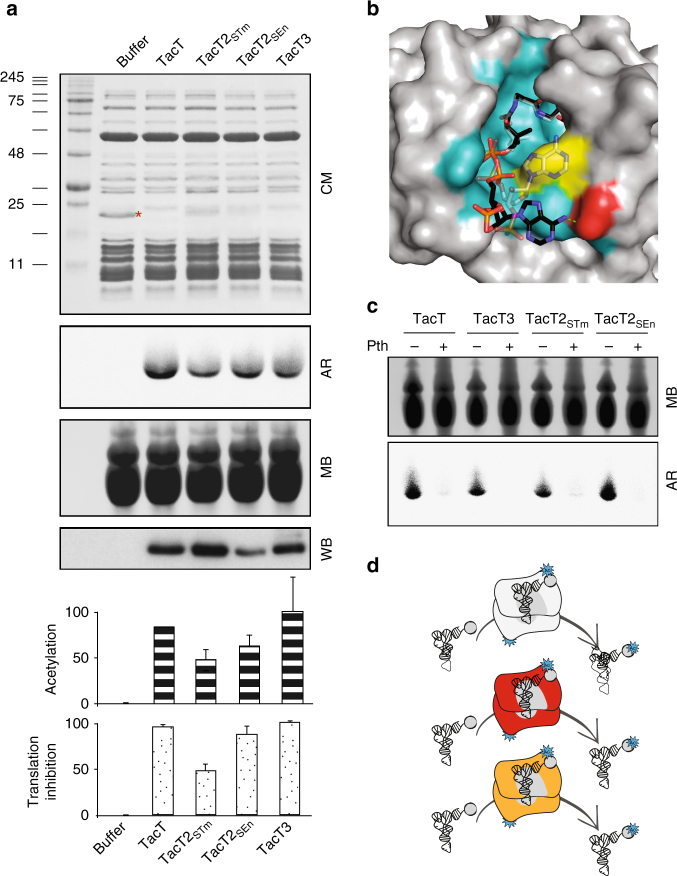
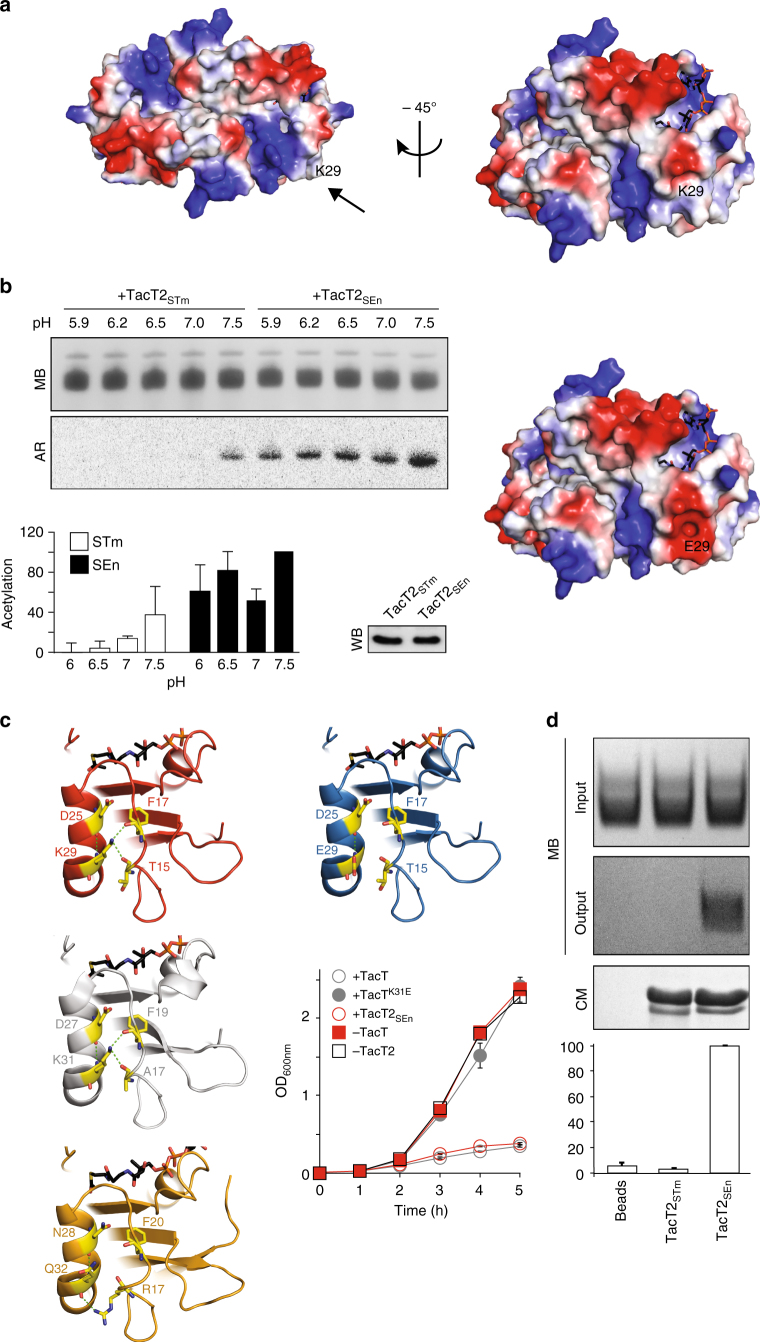
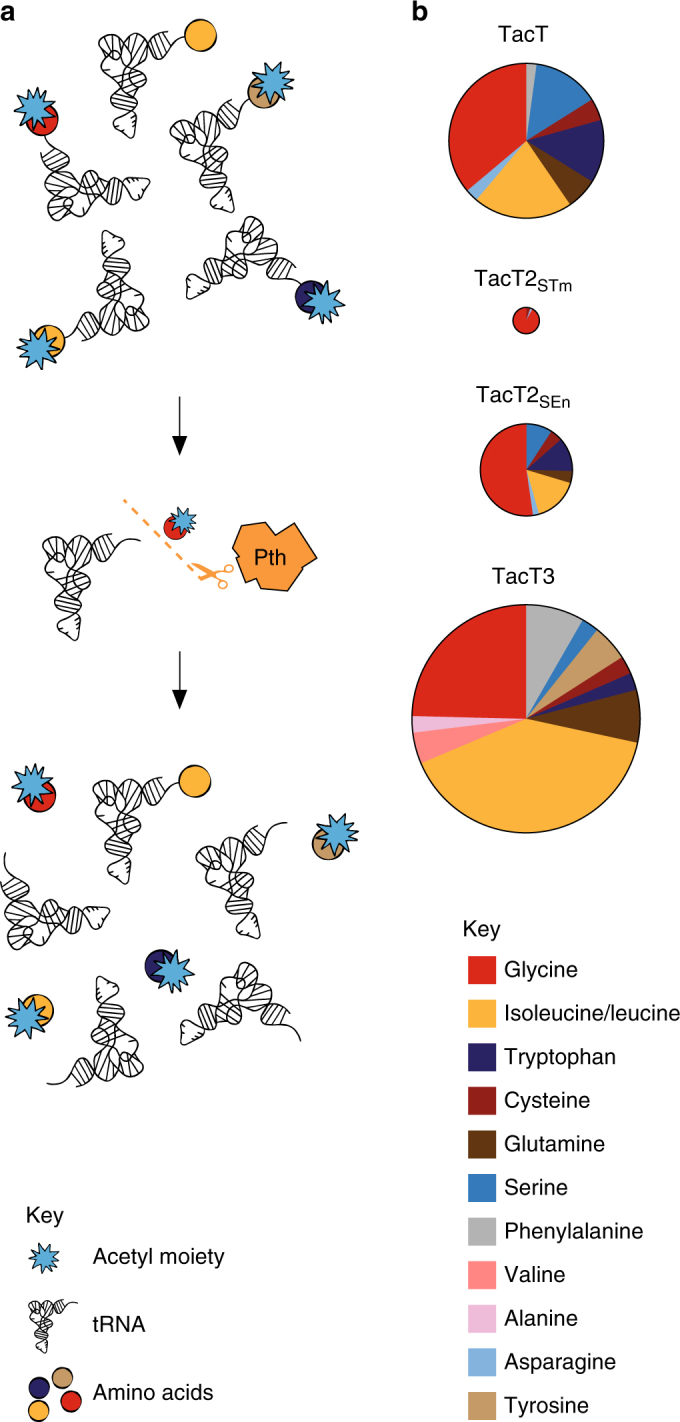
Non-typhoidal Salmonella strains are responsible for invasive infections associated with high mortality and recurrence in sub-Saharan Africa, and there is strong evidence for clonal relapse following antibiotic treatment. Persisters are non-growing bacteria that are thought to be responsible for the recalcitrance of many infections to antibiotics. Toxin-antitoxin systems are stress-responsive elements that are important for Salmonella persister formation, specifically during infection. Here, we report the analysis of persister formation of clinical invasive strains of Salmonella Typhimurium and Enteritidis in human primary macrophages. We show that all the invasive clinical isolates of both serovars that we tested produce high levels of persisters following internalization by human macrophages. Our genome comparison reveals that S. Enteritidis and S. Typhimurium strains contain three acetyltransferase toxins that we characterize structurally and functionally. We show that all induce the persister state by inhibiting translation through acetylation of aminoacyl-tRNAs. However, they differ in their potency and target partially different subsets of aminoacyl-tRNAs, potentially accounting for their non-redundant effect.
Conflict of interest statement
The authors declare no competing interests.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical