Yersinia pestis Survival and Replication in Potential Ameba Reservoir
- PMID: 29350155
- PMCID: PMC5782900
- DOI: 10.3201/eid2402.171065
Yersinia pestis Survival and Replication in Potential Ameba Reservoir
Abstract
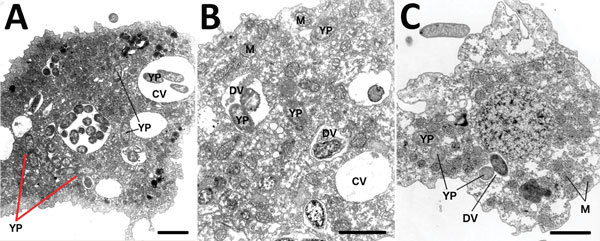
Plague ecology is characterized by sporadic epizootics, then periods of dormancy. Building evidence suggests environmentally ubiquitous amebae act as feral macrophages and hosts to many intracellular pathogens. We conducted environmental genetic surveys and laboratory co-culture infection experiments to assess whether plague bacteria were resistant to digestion by 5 environmental ameba species. First, we demonstrated that Yersinia pestis is resistant or transiently resistant to various ameba species. Second, we showed that Y. pestis survives and replicates intracellularly within Dictyostelium discoideum amebae for ˃48 hours postinfection, whereas control bacteria were destroyed in <1 hour. Finally, we found that Y. pestis resides within ameba structures synonymous with those found in infected human macrophages, for which Y. pestis is a competent pathogen. Evidence supporting amebae as potential plague reservoirs stresses the importance of recognizing pathogen-harboring amebae as threats to public health, agriculture, conservation, and biodefense.
Keywords: Dictyostelium discoideum; Plague; Yersinia pestis; ameba; amoeba; amoebae; biological evolution; coccobacillus; disease reservoir; ecology; endemic; epizootic; eukaryote; feral macrophage; phagocytosis; slime mold; trophozoite.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Bertherat E; World Health Organization. Plague around the world, 2010–2015. Wkly Epidemiol Rec. 2016;91:89–93. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases