Leishmania mortality in sand fly blood meal is not species-specific and does not result from direct effect of proteinases
- PMID: 29335002
- PMCID: PMC5769529
- DOI: 10.1186/s13071-018-2613-2
Leishmania mortality in sand fly blood meal is not species-specific and does not result from direct effect of proteinases
Abstract
Background: Leishmania development in sand flies is confined to the alimentary tract and is closely connected with blood meal digestion. Previously, it has been published that activities of sand fly midgut proteases are harmful to Leishmania, especially to amastigote-promastigote transition forms. However, our experiments with various Leishmania-sand fly pairs gave quite opposite results.
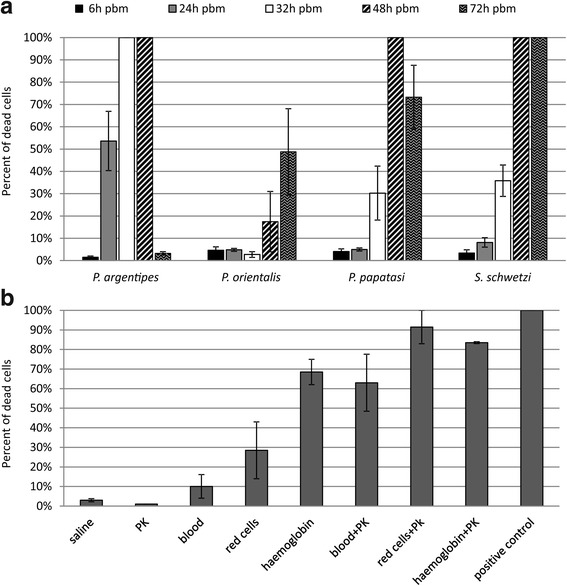
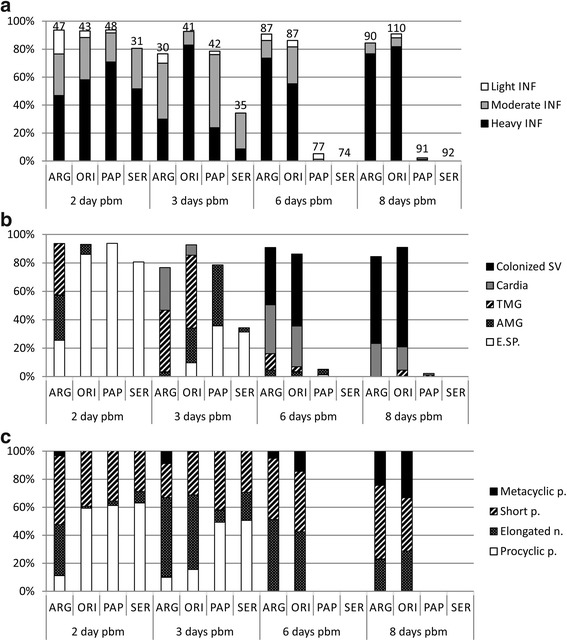
Methods: We evaluated the effect of semi-digested midgut content on different life stages of Leishmania donovani and Leishmania major in vitro. Various morphological forms of parasites, including macrophage-derived amastigotes and transition forms, were incubated 2 h with midguts dissected at various intervals (6-72 h) post-blood meal or with commercially available proteinase, and their viability was determined using flow cytometry. In parallel, using amastigote-initiated experimental infections, we compared development of L. donovani in sand flies that are either susceptible (Phlebotomus argentipes and P. orientalis) or refractory (P. papatasi and Sergentomyia schwetzi) to this parasite.
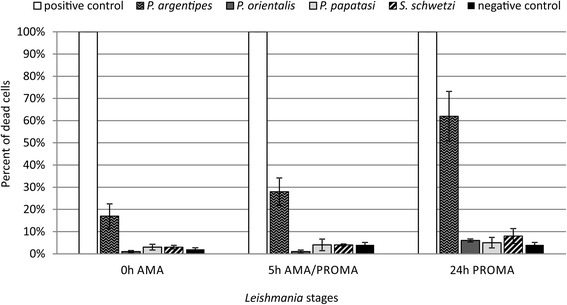
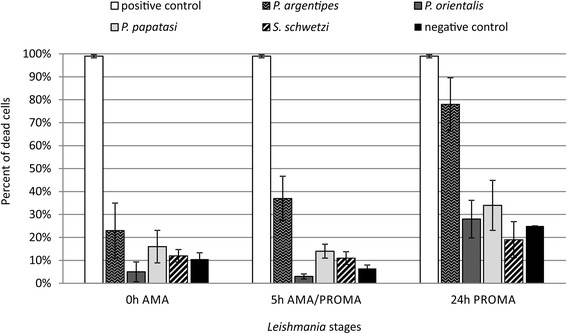
Results: In vitro, sand fly midgut homogenates affected L. major and L. donovani in a similar way; in all sand fly species, the most significant mortality effect was observed by the end of the blood meal digestion process. Surprisingly, the most susceptible Leishmania stages were promastigotes, while mortality of transforming parasites and amastigotes was significantly lower. Parasites were also susceptible to killing by rabbit blood in combination with proteinase, but resistant to proteinase itself. In vivo, L. donovani developed late-stage infections in both natural vectors; in P. argentipes the development was much faster than in P. orientalis. On the other hand, in refractory species P. papatasi and S. schwetzi, promastigotes survived activity of digestive enzymes but were lost during defecation.
Conclusions: We demonstrated that Leishmania transition forms are more resistant to the killing effect of semi-digested blood meal than 24 h-old promastigotes. Data suggest that Leishmania mortality is not caused directly by sand fly proteases, we assume that this mortality results from toxic products of blood meal digestion. Survival of L. donovani promastigotes in refractory sand flies until blood meal defecation, together with similar mortality of Leishmania parasites incubated in vitro with midgut homogenates of susceptible as well as refractory species, contradict the previously raised hypotheses about the role of midgut proteases in sand fly vector competence to Leishmania.
Keywords: Blood meal digestion; Leishmania donovani; Leishmania major; Phlebotomus; Proteases; Sand fly; Sergentomyia.
Conflict of interest statement
Ethics approval
Animals used for maintenance of sand flies colonies were maintained and handled in the animal facility of Charles University in Prague in accordance with institutional guidelines and Czech legislation (Act No. 246/1992 and 359/2012 coll. on Protection of Animals against Cruelty in present statutes at large), which complies with all relevant EU guidelines for experimental animals. All experiments were approved by the Committee on the Ethics of Laboratory Experiments of the Charles University in Prague and were performed under the Certificate of Competency (Registration Number: CZ 03069).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical