Genome wide comparison of Ethiopian Leishmania donovani strains reveals differences potentially related to parasite survival
- PMID: 29315303
- PMCID: PMC5777657
- DOI: 10.1371/journal.pgen.1007133
Genome wide comparison of Ethiopian Leishmania donovani strains reveals differences potentially related to parasite survival
Abstract
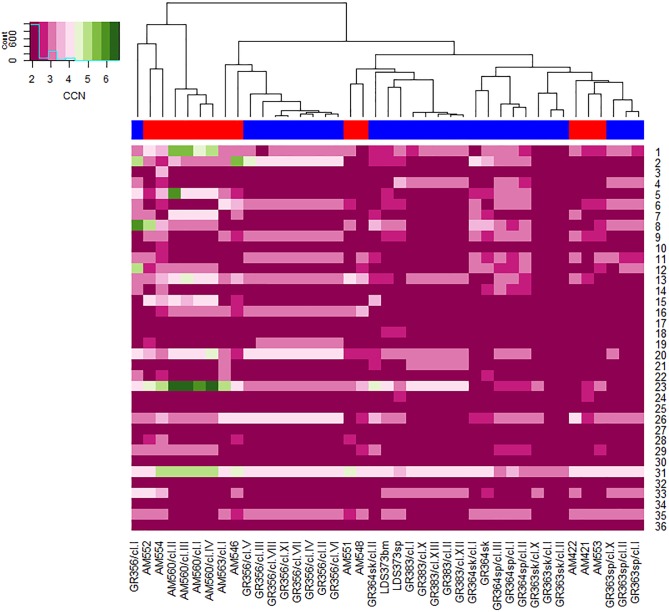
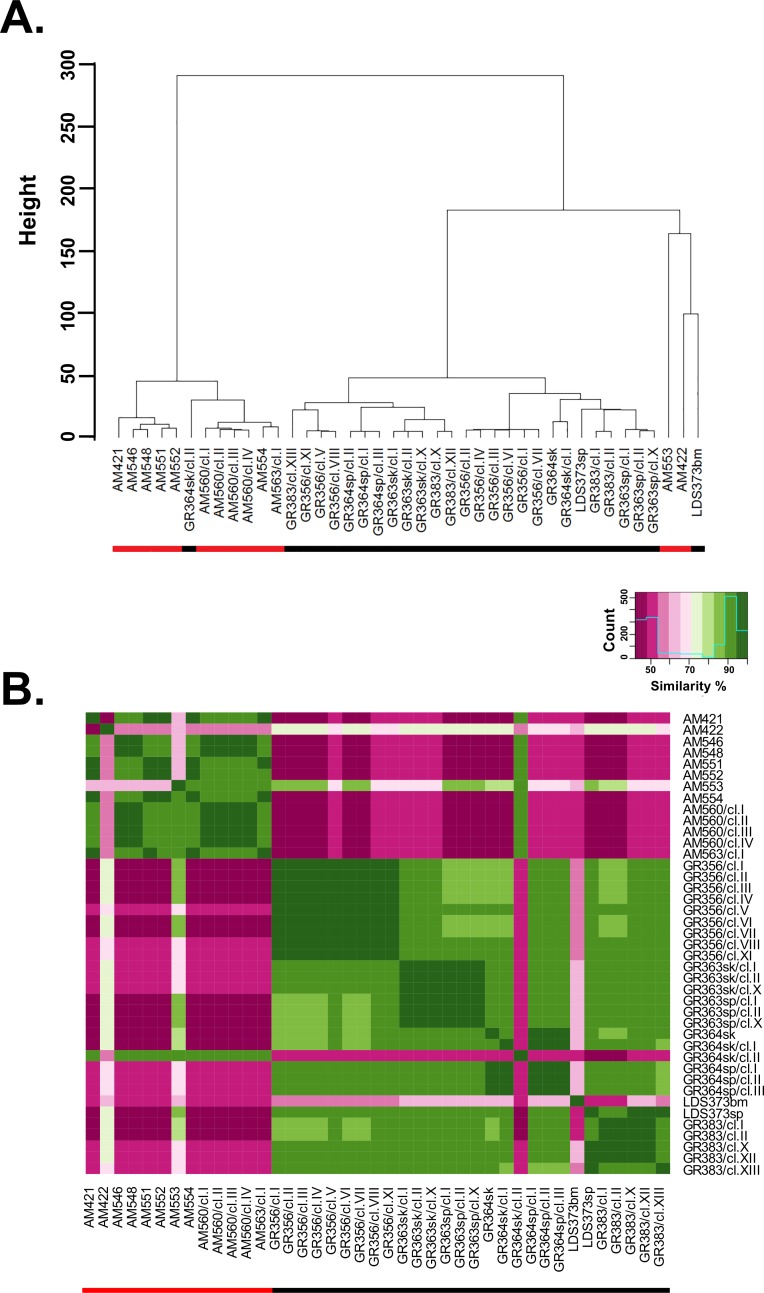
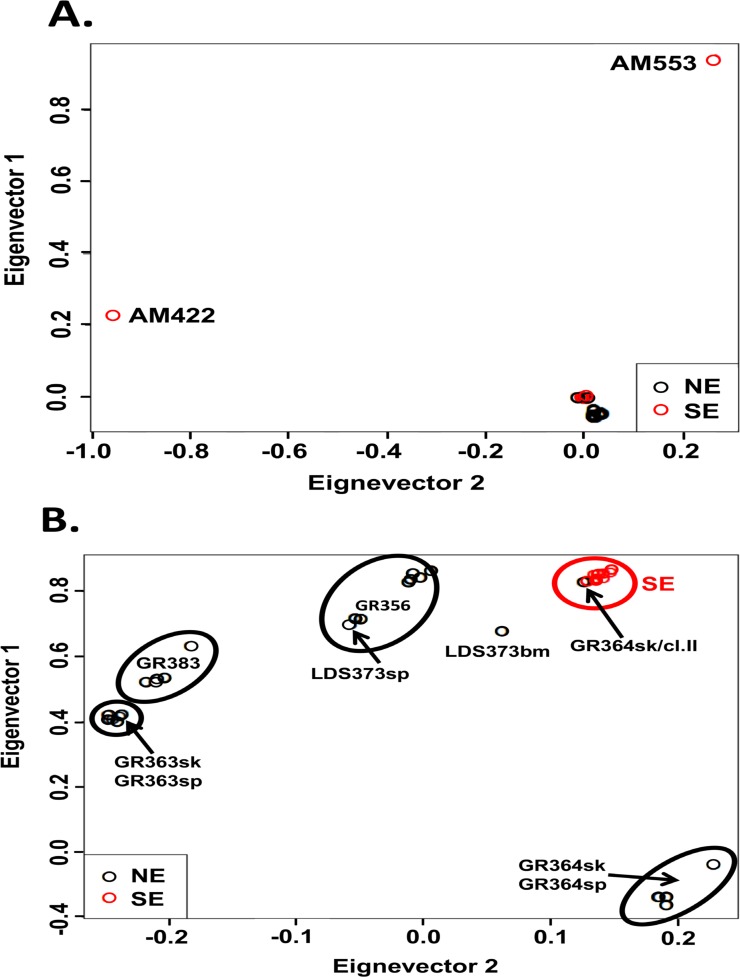
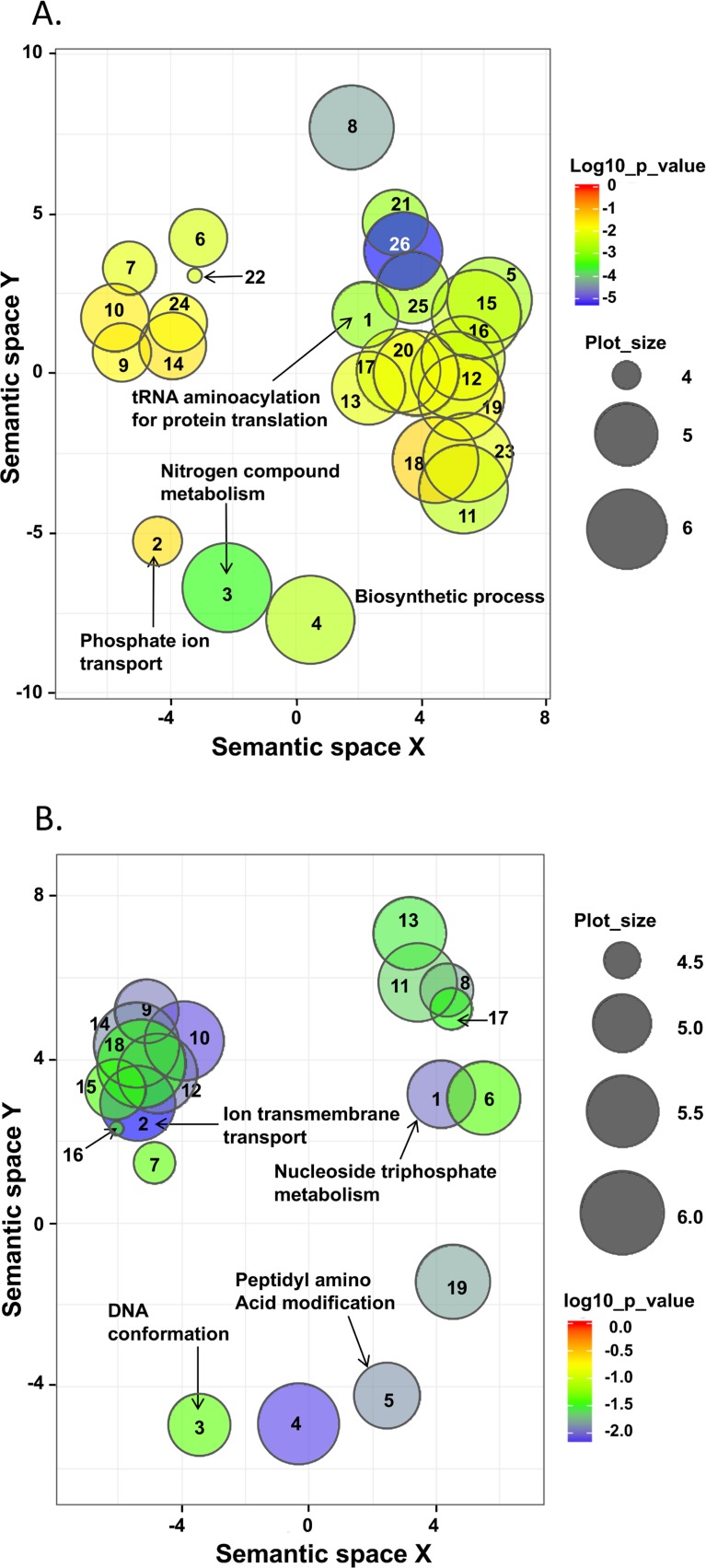
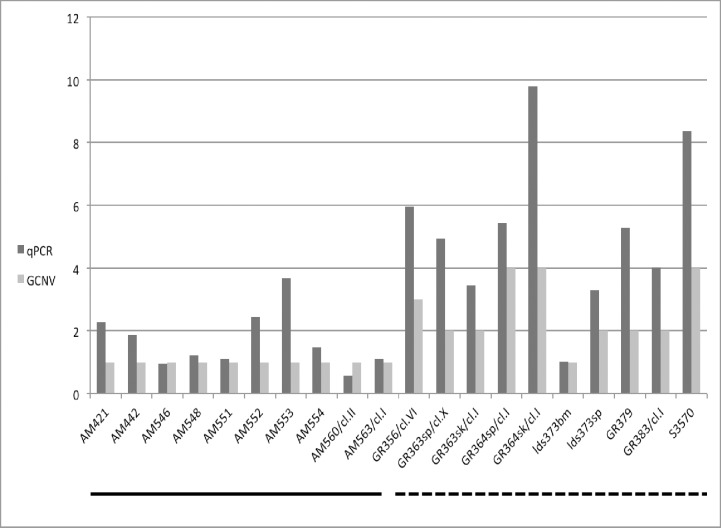
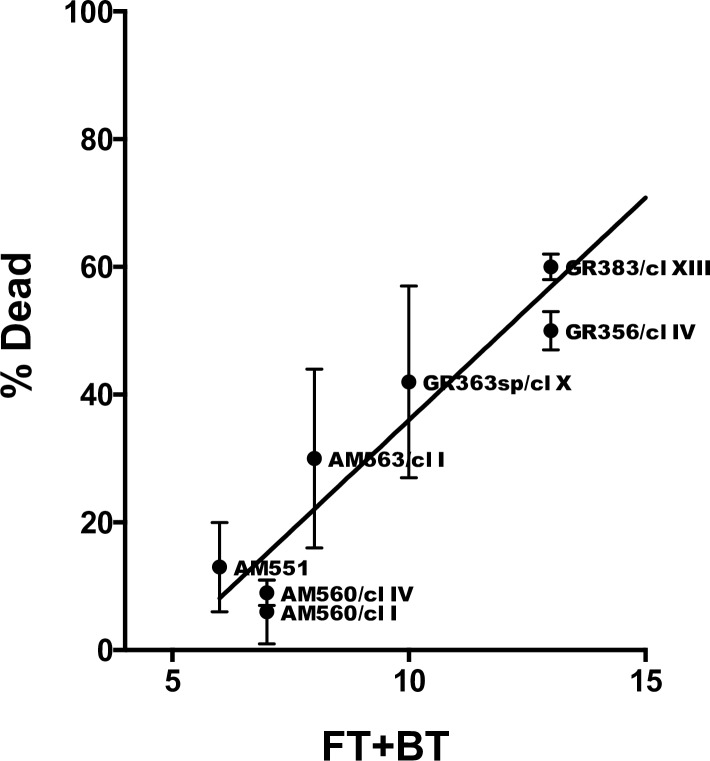
Leishmania donovani is the main cause of visceral leishmaniasis (VL) in East Africa. Differences between northern Ethiopia/Sudan (NE) and southern Ethiopia (SE) in ecology, vectors, and patient sensitivity to drug treatment have been described, however the relationship between differences in parasite genotype between these two foci and phenotype is unknown. Whole genomic sequencing (WGS) was carried out for 41 L. donovani strains and clones from VL and VL/HIV co-infected patients in NE (n = 28) and SE (n = 13). Chromosome aneuploidy was observed in all parasites examined with each isolate exhibiting a unique karyotype. Differences in chromosome ploidy or karyotype were not correlated with the geographic origin of the parasites. However, correlation between single nucleotide polymorphism (SNP) and geographic origin was seen for 38/41 isolates, separating the NE and SE parasites into two large groups. SNP restricted to NE and SE groups were associated with genes involved in viability and parasite resistance to drugs. Unique copy number variation (CNV) were also associated with NE and SE parasites, respectively. One striking example is the folate transporter (FT) family genes (LdBPK_100390, LdBPK_100400 and LdBPK_100410) on chromosome 10 that are single copy in all 13 SE isolates, but either double copy or higher in 39/41 NE isolates (copy number 2-4). High copy number (= 4) was also found for one Sudanese strain examined. This was confirmed by quantitative polymerase chain reaction for LdBPK_100400, the L. donovani FT1 transporter homolog. Good correlation (p = 0.005) between FT copy number and resistance to methotrexate (0.5 mg/ml MTX) was also observed with the haploid SE strains examined showing higher viability than the NE strains at this concentration. Our results emphasize the advantages of whole genome analysis to shed light on vital parasite processes in Leishmania.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Alvar J, Velez ID, Bern C, Herrero M, Desjeux P, Cano J, et al. Leishmaniasis worldwide and global estimates of its incidence. PLoS One. 2012;7(5):e35671 doi: 10.1371/journal.pone.0035671 - DOI - PMC - PubMed
-
- Alvar J, Aparicio P, Aseffa A, Den Boer M, Canavate C, Dedet JP, et al. The relationship between leishmaniasis and AIDS: the second 10 years. Clin Microbiol Rev. 2008;21(2):334–59. doi: 10.1128/CMR.00061-07 - DOI - PMC - PubMed
-
- Hurissa Z, Gebre-Silassie S, Hailu W, Tefera T, Lalloo DG, Cuevas LE, et al. Clinical characteristics and treatment outcome of patients with visceral leishmaniasis and HIV co-infection in northwest Ethiopia. Trop Med Int Health. 2010;15(7):848–55. doi: 10.1111/j.1365-3156.2010.02550.x - DOI - PubMed
-
- Sundar S, Chakravarty J. Leishmaniasis: an update of current pharmacotherapy. Exp Opin Pharm. 2013;14(1):53–63. doi: 10.1517/14656566.2013.755515 . - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases