In situ immune response and mechanisms of cell damage in central nervous system of fatal cases microcephaly by Zika virus
- PMID: 29311619
- PMCID: PMC5758755
- DOI: 10.1038/s41598-017-17765-5
In situ immune response and mechanisms of cell damage in central nervous system of fatal cases microcephaly by Zika virus
Abstract
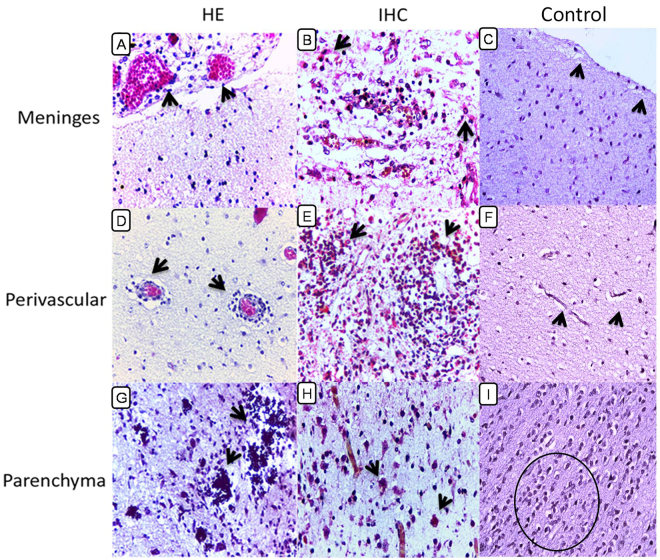
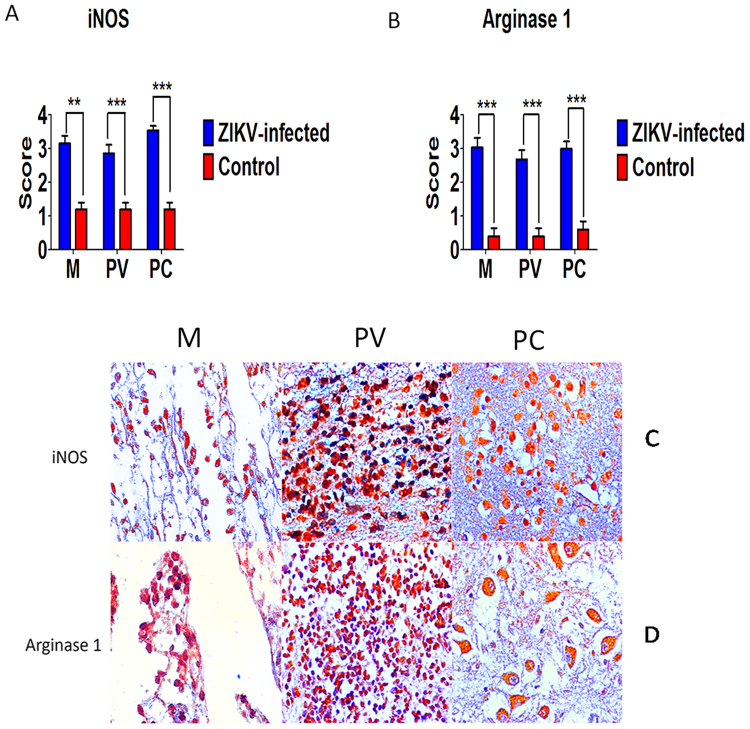
Zika virus (ZIKV) has recently caused a pandemic disease, and many cases of ZIKV infection in pregnant women resulted in abortion, stillbirth, deaths and congenital defects including microcephaly, which now has been proposed as ZIKV congenital syndrome. This study aimed to investigate the in situ immune response profile and mechanisms of neuronal cell damage in fatal Zika microcephaly cases. Brain tissue samples were collected from 15 cases, including 10 microcephalic ZIKV-positive neonates with fatal outcome and five neonatal control flavivirus-negative neonates that died due to other causes, but with preserved central nervous system (CNS) architecture. In microcephaly cases, the histopathological features of the tissue samples were characterized in three CNS areas (meninges, perivascular space, and parenchyma). The changes found were mainly calcification, necrosis, neuronophagy, gliosis, microglial nodules, and inflammatory infiltration of mononuclear cells. The in situ immune response against ZIKV in the CNS of newborns is complex. Despite the predominant expression of Th2 cytokines, other cytokines such as Th1, Th17, Treg, Th9, and Th22 are involved to a lesser extent, but are still likely to participate in the immunopathogenic mechanisms of neural disease in fatal cases of microcephaly caused by ZIKV.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Barzon, L., Trevisan, M., Sinigaglia, A., Lavezzo, E. & Palù, G. Zika virus pathogenesis to disease control. FEMS Microbiol Lett. 363, 10.1093/femsle/fnw202 (2016). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical