Viral and cellular N6-methyladenosine and N6,2'-O-dimethyladenosine epitranscriptomes in the KSHV life cycle
- PMID: 29109479
- PMCID: PMC6138870
- DOI: 10.1038/s41564-017-0056-8
Viral and cellular N6-methyladenosine and N6,2'-O-dimethyladenosine epitranscriptomes in the KSHV life cycle
Abstract
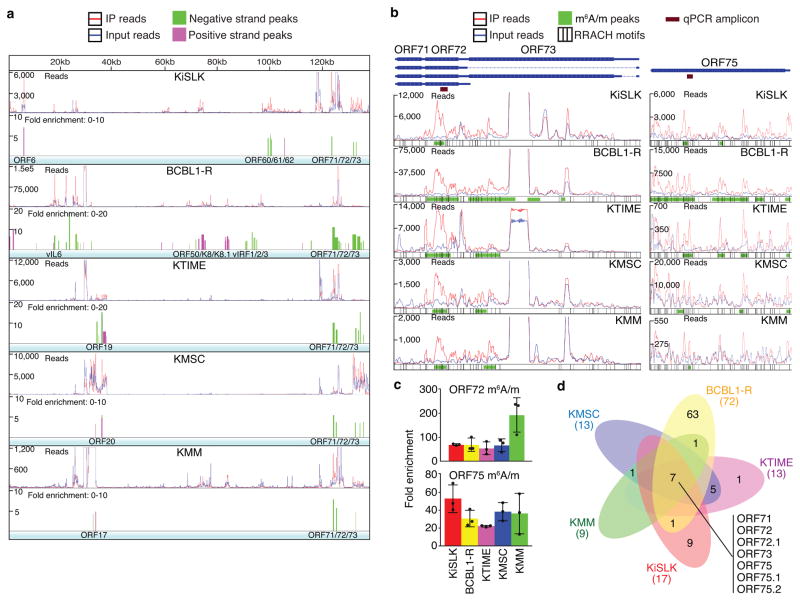
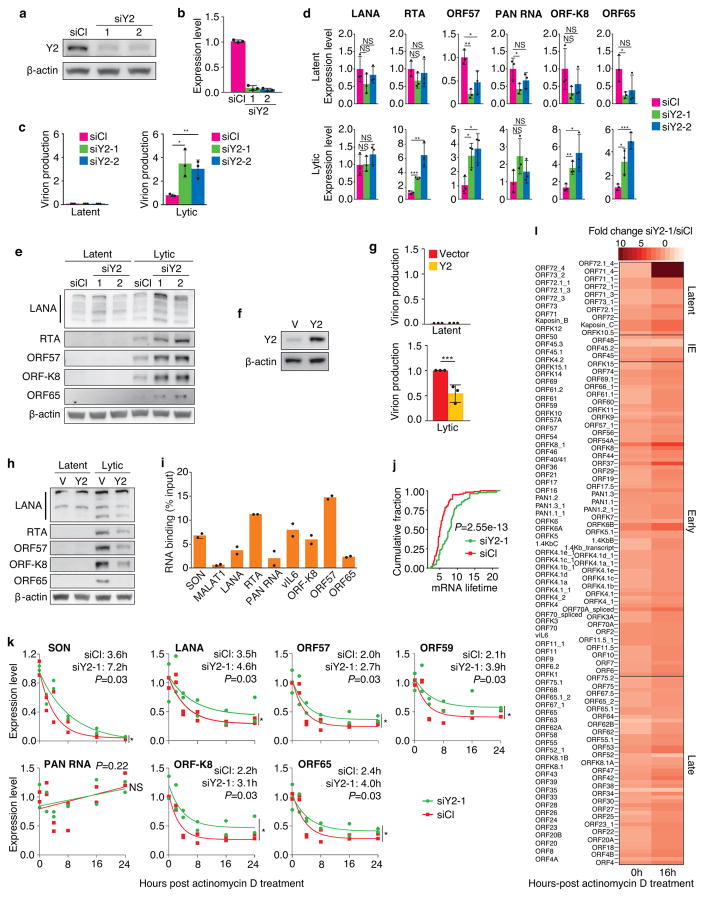
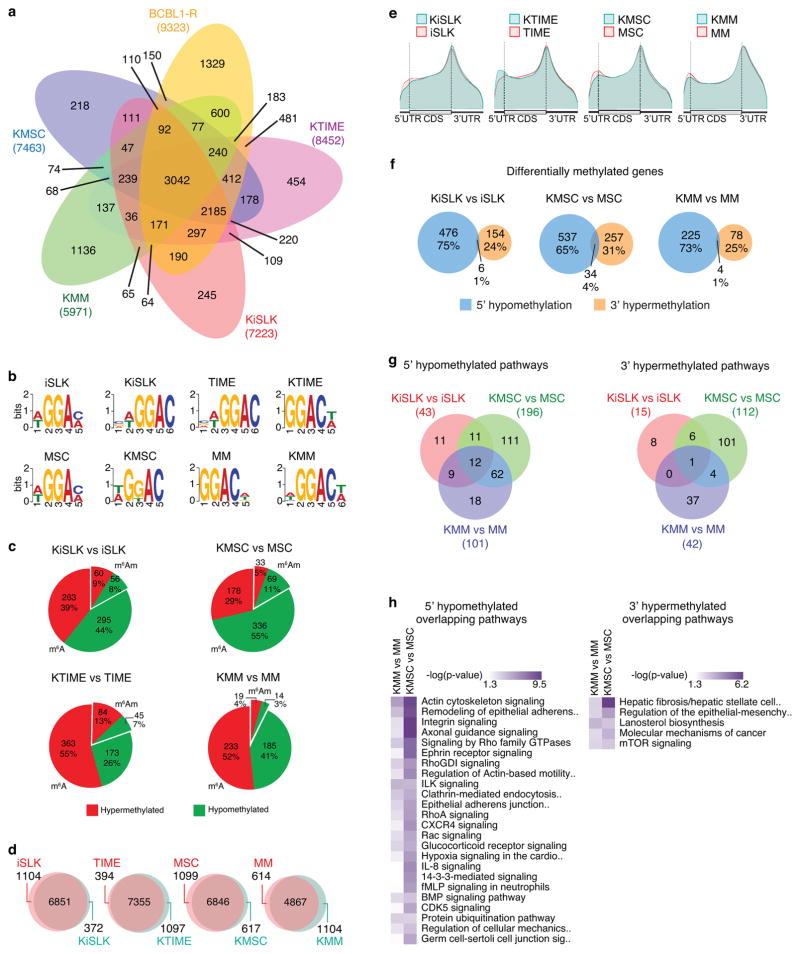
N6-methyladenosine (m6A) and N6,2'-O-dimethyladenosine (m6Am) modifications (m6A/m) of messenger RNA mediate diverse cellular functions. Oncogenic Kaposi's sarcoma-associated herpesvirus (KSHV) has latent and lytic replication phases that are essential for the development of KSHV-associated cancers. To date, the role of m6A/m in KSHV replication and tumorigenesis is unclear. Here, we provide mechanistic insights by examining the viral and cellular m6A/m epitranscriptomes during KSHV latent and lytic infection. KSHV transcripts contain abundant m6A/m modifications during latent and lytic replication, and these modifications are highly conserved among different cell types and infection systems. Knockdown of YTHDF2 enhanced lytic replication by impeding KSHV RNA degradation. YTHDF2 binds to viral transcripts and differentially mediates their stability. KSHV latent infection induces 5' untranslated region (UTR) hypomethylation and 3'UTR hypermethylation of the cellular epitranscriptome, regulating oncogenic and epithelial-mesenchymal transition pathways. KSHV lytic replication induces dynamic reprogramming of epitranscriptome, regulating pathways that control lytic replication. These results reveal a critical role of m6A/m modifications in KSHV lifecycle and provide rich resources for future investigations.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases