New insights into experimental visceral leishmaniasis: Real-time in vivo imaging of Leishmania donovani virulence
- PMID: 28945751
- PMCID: PMC5629011
- DOI: 10.1371/journal.pntd.0005924
New insights into experimental visceral leishmaniasis: Real-time in vivo imaging of Leishmania donovani virulence
Abstract
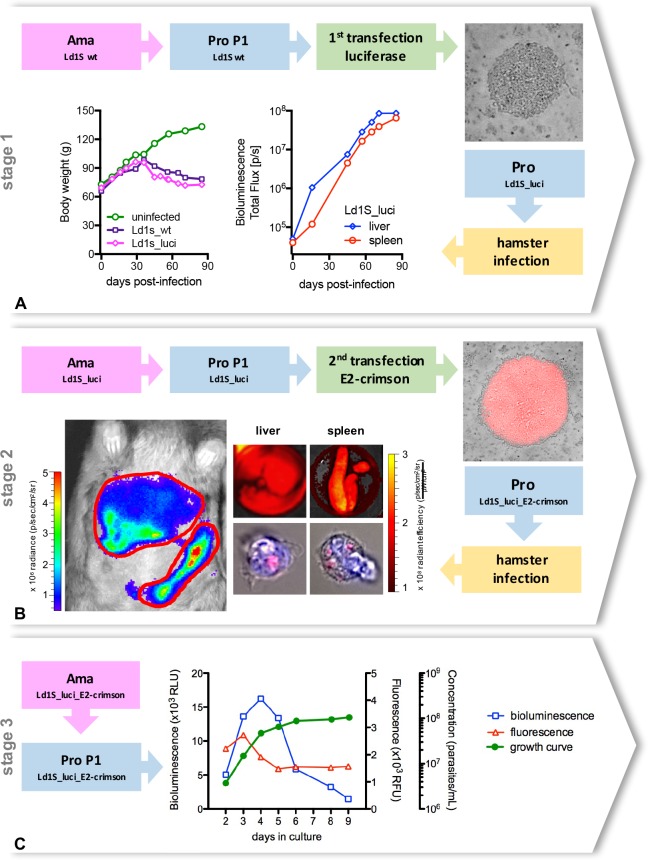
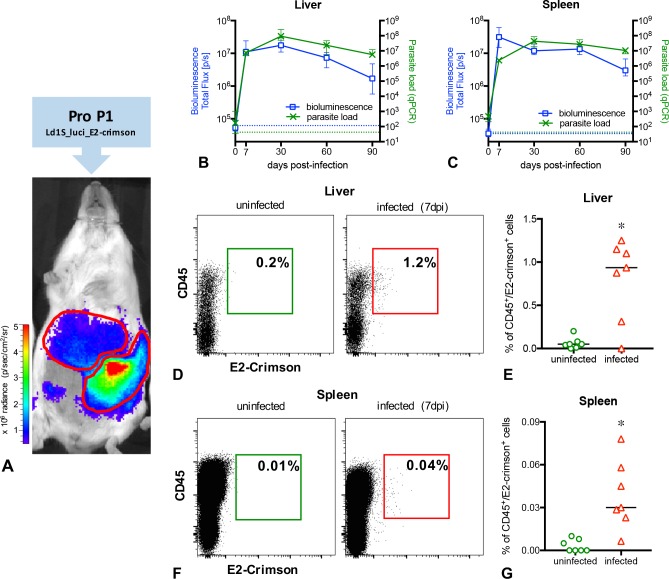
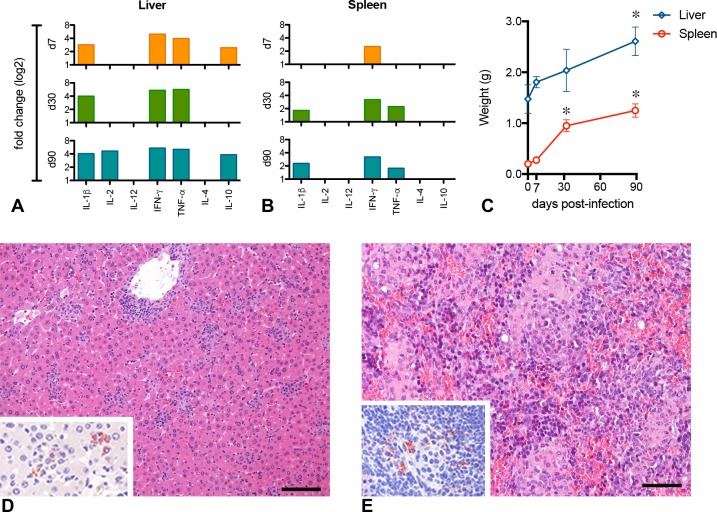
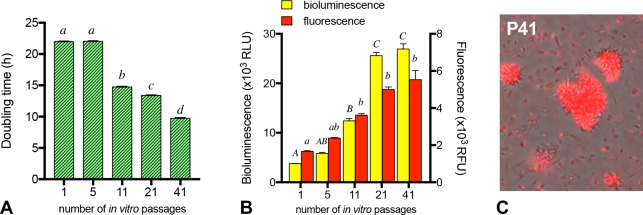
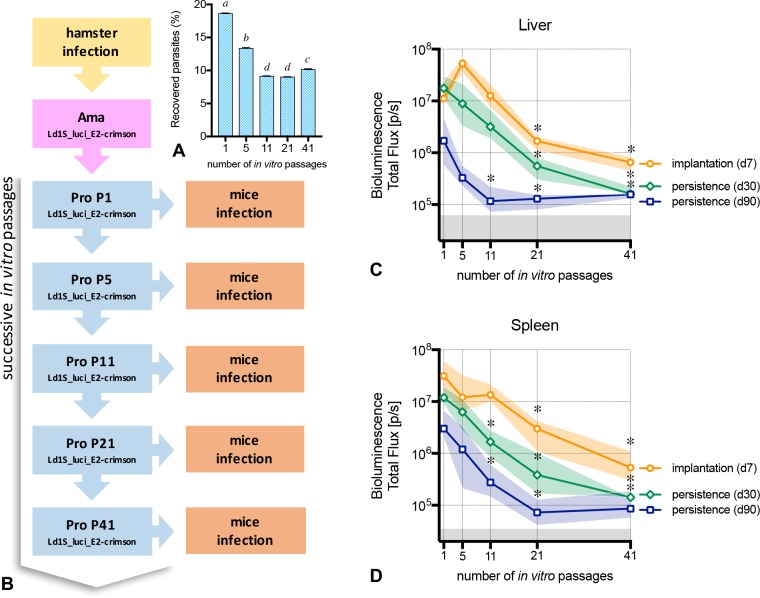
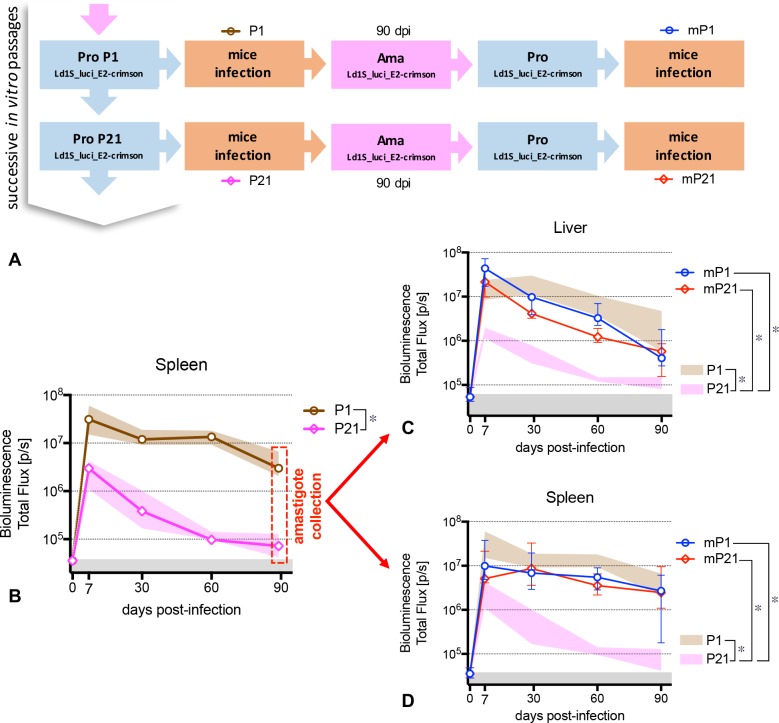
Visceral leishmaniasis is an insidious neglected disease with worldwide distribution. It is caused by parasites from the Leishmania donovani complex, which are able to be transmitted by different species of phlebotomine sand flies and to infect numerous mammal hosts. Despite the high number of people infected or at risk, and the remarkable quantity of studies focusing on this disease, a proper experimental model to efficiently decipher the infectious process of visceral leishmaniasis taking into account the nuances of parasite’s virulence and the duration of the infection is still lacking. Therefore, using golden Syrian hamsters and BALB/c mice, state-of-the-art genetic manipulation applied on a fully virulent L. donovani strain and in vivo imaging approaches, we describe herein three benefits for experimental visceral leishmaniasis: (i) the development of a double transfected bioluminescent (firefly luciferase) and fluorescent (E2-crimson) virulent strain of L. donovani (Ld1S_luci_E2-crimson), favoring a wide range of both in vivo and in vitro investigations, (ii) the establishment of a non-invasive mouse model to evaluate the infectious process during visceral leishmaniasis and the parasite’s virulence in real time, allowing longitudinal studies with the same animals, and (iii) the elaboration of a suitable method to reinstate (and verify anew) the virulence in a population of attenuated parasites, by recovering persistent parasites from chronic infected mice. Consequently, these results open up new perspectives on the study of visceral leishmaniasis, especially in the fields of therapeutics and vaccinology, since the model described herein renders now possible long-lasting follow up studies, with easy and accurate day-by-day verifications of the infection status along with a reduced number of laboratory animals.
Trial registration: ClinicalTrials.gov 2013-0047.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- WHO (2010) Control of the leishmaniasis: report of a meeting of the WHO Expert Committee on the Control of Leishmaniases, Geneva, 22–26 March 2010. WHO Technical Report No. 949.
-
- Alvar J, Vélez ID, Bern C, Herrero M, Desjeux P, et al. (2012) Leishmaniasis Worldwide and Global Estimates of Its Incidence. PLoS ONE 7: e35671 doi: 10.1371/journal.pone.0035671 - DOI - PMC - PubMed
-
- Baneth G, Koutinas AF, Solano-Gallego L, Bourdeau P, Ferrer L (2008) Canine leishmaniosis–new concepts and insights on an expanding zoonosis: part one. Trends in Parasitology 24: 324–330. doi: 10.1016/j.pt.200804001 - DOI - PubMed
-
- Fernández-Cotrina J, Iniesta V, Belinchón-Lorenzo S, Muñoz-Madrid R, Serrano F, et al. (2013) Experimental model for reproduction of canine visceral leishmaniosis by Leishmania infantum. Veterinary Parasitology 192: 118–128. doi: 10.1016/j.vetpar.201210002 - DOI - PubMed
-
- Melby PC, Chandrasekar B, Zhao W, Coe JE (2001) The Hamster as a Model of Human Visceral Leishmaniasis: Progressive Disease and Impaired Generation of Nitric Oxide in the Face of a Prominent Th1-Like Cytokine Response. The Journal of Immunology 166: 1912–1920. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources