A Review and Database of Snake Venom Proteomes
- PMID: 28927001
- PMCID: PMC5618223
- DOI: 10.3390/toxins9090290
A Review and Database of Snake Venom Proteomes
Abstract
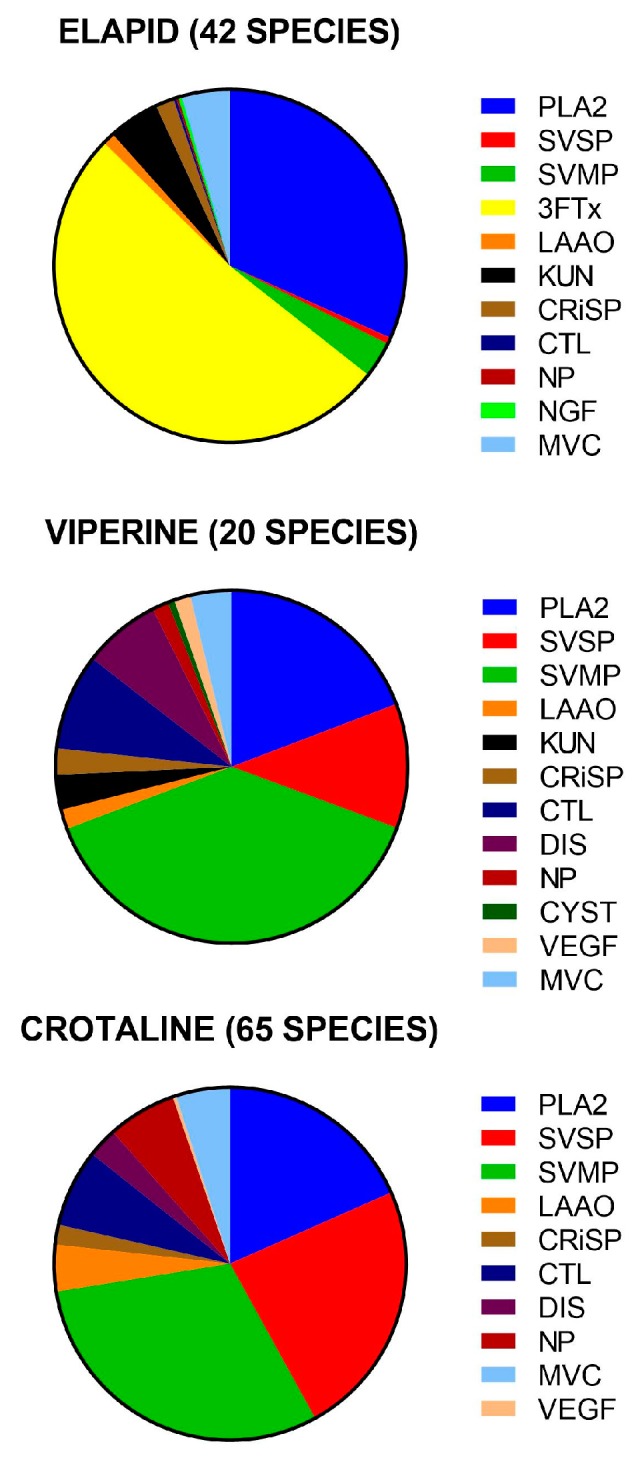
Advances in the last decade combining transcriptomics with established proteomics methods have made possible rapid identification and quantification of protein families in snake venoms. Although over 100 studies have been published, the value of this information is increased when it is collated, allowing rapid assimilation and evaluation of evolutionary trends, geographical variation, and possible medical implications. This review brings together all compositional studies of snake venom proteomes published in the last decade. Compositional studies were identified for 132 snake species: 42 from 360 (12%) Elapidae (elapids), 20 from 101 (20%) Viperinae (true vipers), 65 from 239 (27%) Crotalinae (pit vipers), and five species of non-front-fanged snakes. Approximately 90% of their total venom composition consisted of eight protein families for elapids, 11 protein families for viperines and ten protein families for crotalines. There were four dominant protein families: phospholipase A2s (the most common across all front-fanged snakes), metalloproteases, serine proteases and three-finger toxins. There were six secondary protein families: cysteine-rich secretory proteins, l-amino acid oxidases, kunitz peptides, C-type lectins/snaclecs, disintegrins and natriuretic peptides. Elapid venoms contained mostly three-finger toxins and phospholipase A2s and viper venoms metalloproteases, phospholipase A2s and serine proteases. Although 63 protein families were identified, more than half were present in <5% of snake species studied and always in low abundance. The importance of these minor component proteins remains unknown.
Keywords: elapid; proteomics; snakes; toxins; venom; viper.
Conflict of interest statement
The authors declare no conflict of interest. Any conclusions drawn are dependent on the accuracy of the original papers.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Jackson T.N.W., Young B., Underwood G., McCarthy C., Kochva E., Vidal N., van der Weerd L., Nabuurs R., Dobson J., Whitehead D., et al. Endless forms most beautiful: The evolution of ophidian oral glands, including the venom system, and the use of appropriate terminology for homologous structures. Zoomorphology. 2017;136:107–130. doi: 10.1007/s00435-016-0332-9. - DOI
-
- Chatrath S.T., Chapeaurouge A., Lin Q., Lim T.K., Dunstan N., Mirtschin P., Kumar P.P., Kini R.M. Identification of Novel Proteins from the Venom of a Cryptic Snake Drysdalia coronoides by a Combined Transcriptomics and Proteomics Approach. J. Proteome Res. 2011;10:739–750. doi: 10.1021/pr1008916. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical