Toxoplasma Co-infection Prevents Th2 Differentiation and Leads to a Helminth-Specific Th1 Response
- PMID: 28791259
- PMCID: PMC5524676
- DOI: 10.3389/fcimb.2017.00341
Toxoplasma Co-infection Prevents Th2 Differentiation and Leads to a Helminth-Specific Th1 Response
Abstract
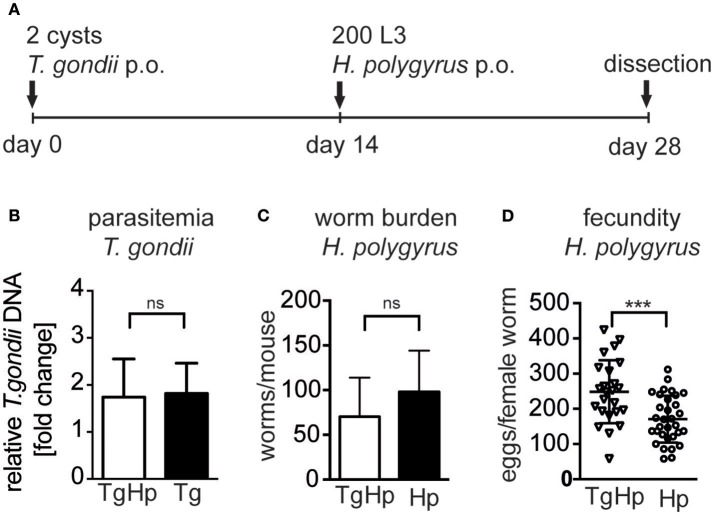
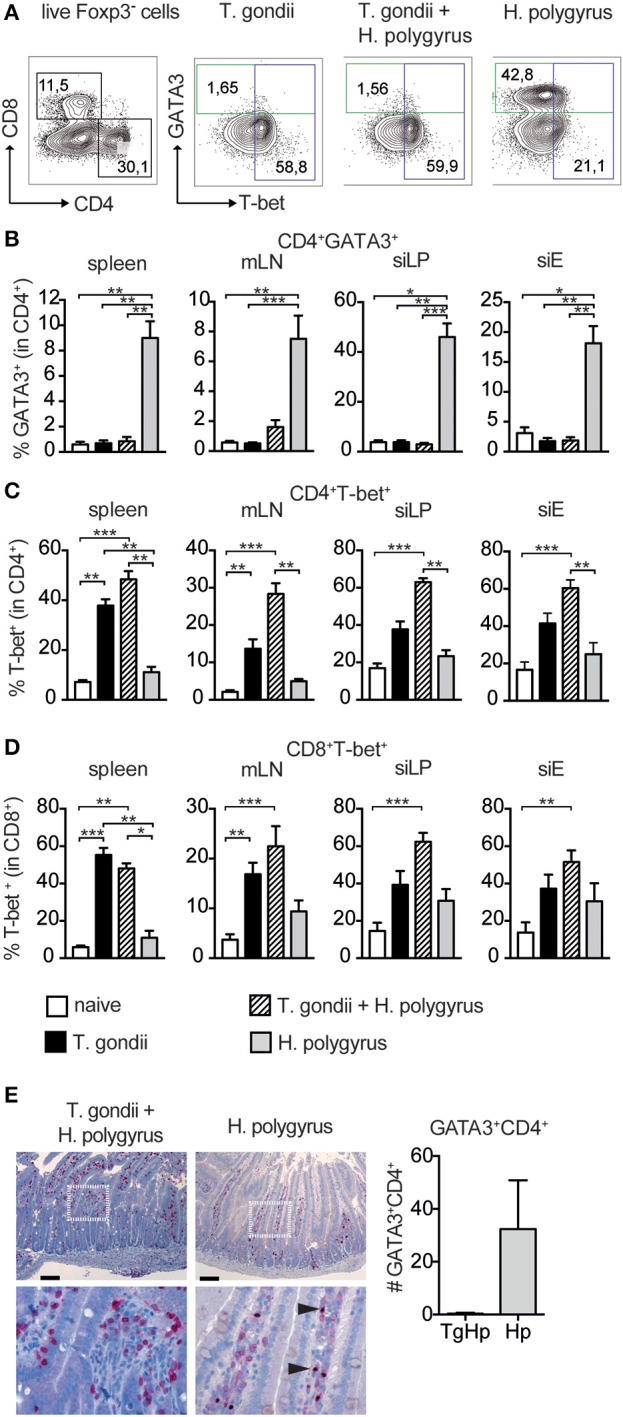
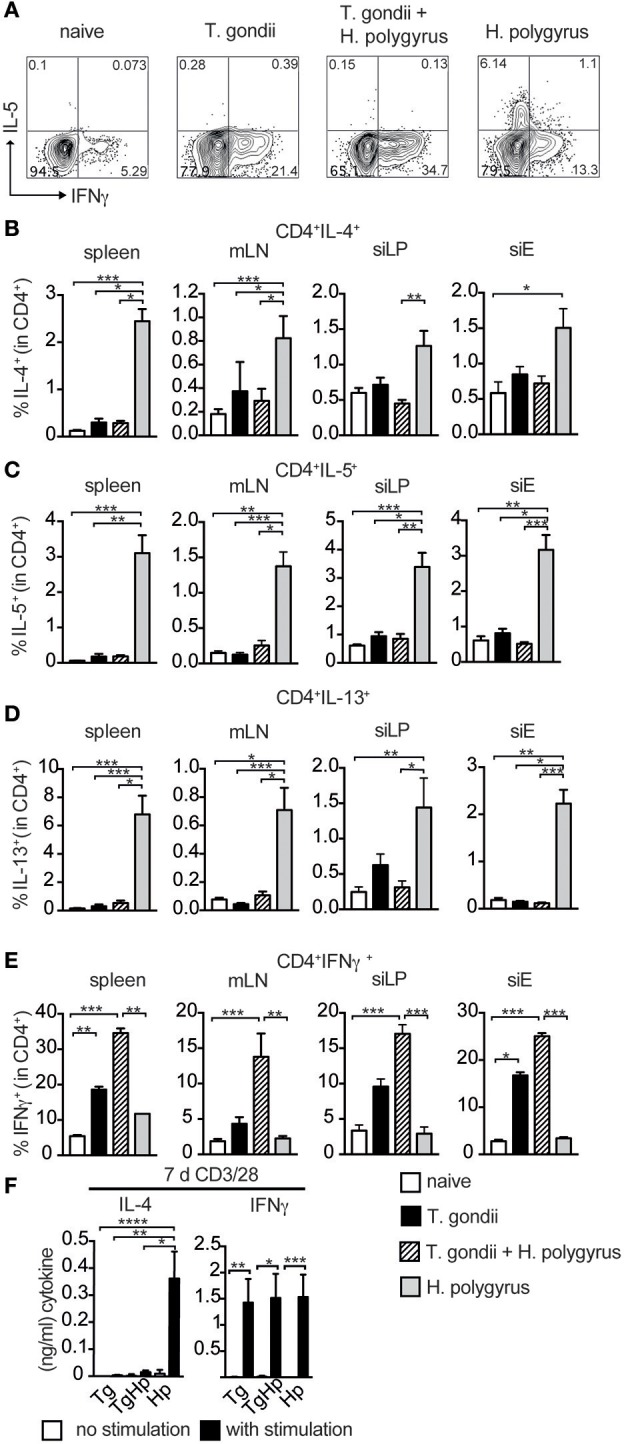
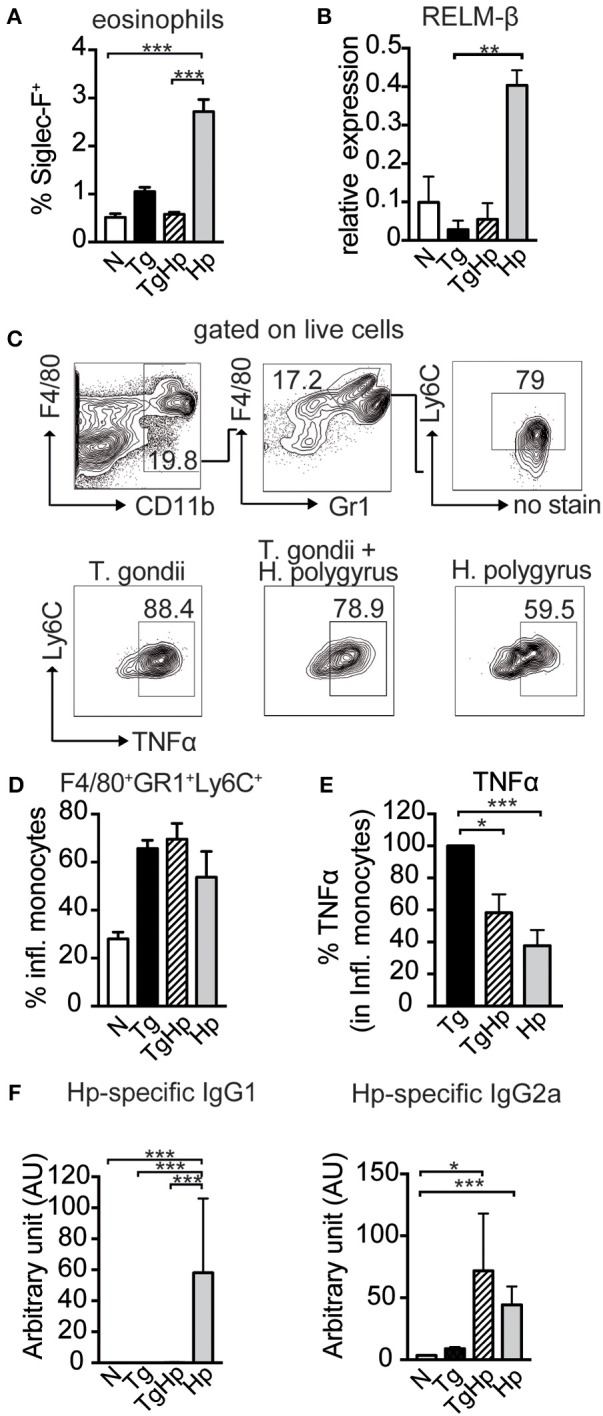
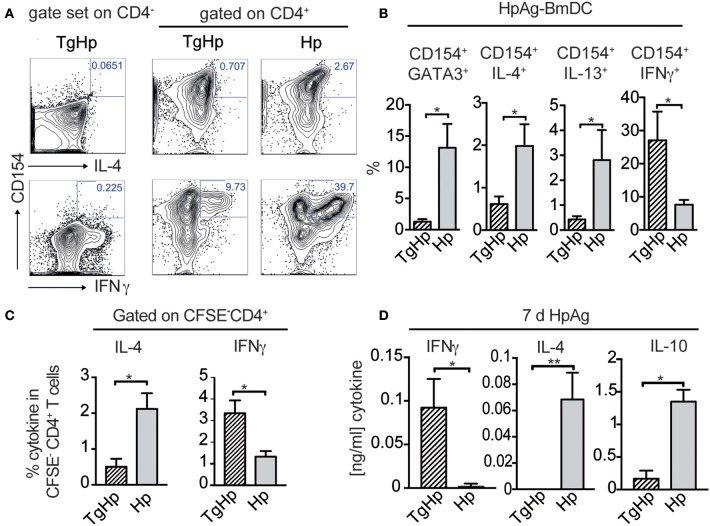
Nematode infections, in particular gastrointestinal nematodes, are widespread and co-infections with other parasites and pathogens are frequently encountered in humans and animals. To decipher the immunological effects of a widespread protozoan infection on the anti-helminth immune response we studied a co-infection with the enteric nematode Heligmosomoides polygyrus in mice previously infected with Toxoplasma gondii. Protective immune responses against nematodes are dependent on parasite-specific Th2 responses associated with IL-4, IL-5, IL-13, IgE, and IgG1 antibodies. In contrast, Toxoplasma gondii infection elicits a strong and protective Th1 immune response characterized by IFN-γ, IL-12, and IgG2a antibodies. Co-infected animals displayed significantly higher worm fecundity although worm burden remained unchanged. In line with this, the Th2 response to H. polygyrus in co-infected animals showed a profound reduction of IL-4, IL-5, IL-13, and GATA-3 expressing T cells. Co-infection also resulted in the lack of eosinophilia and reduced expression of the Th2 effector molecule RELM-β in intestinal tissue. In contrast, the Th1 response to the protozoan parasite was not diminished and parasitemia of T. gondii was unaffected by concurrent helminth infection. Importantly, H. polygyrus specific restimulation of splenocytes revealed H. polygyrus-reactive CD4+ T cells that produce a significant amount of IFN-γ in co-infected animals. This was not observed in animals infected with the nematode alone. Increased levels of H. polygyrus-specific IgG2a antibodies in co-infected mice mirrored this finding. This study suggests that polarization rather than priming of naive CD4+ T cells is disturbed in mice previously infected with T. gondii. In conclusion, a previous T. gondii infection limits a helminth-specific Th2 immune response while promoting a shift toward a Th1-type immune response.
Keywords: Heligmosomoides polygyrus; Th1; Th2; Toxoplasma gondii; co-infection; helminth.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Bahia-Oliveira L. M., da Silva J. A., Peixoto-Rangel A. L., Boechat M. S. B., Oliveira A. M. W. A., Massara C. L., et al. . (2009). Host immune response to Toxoplasma gondii and Ascaris lumbricoides in a highly endemic area: evidence of parasite co-immunomodulation properties influencing the outcome of both infections. Mem. Inst. Oswaldo Cruz 104, 273–280. 10.1590/S0074-02762009000200021 - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials