Covalently linked dengue virus envelope glycoprotein dimers reduce exposure of the immunodominant fusion loop epitope
- PMID: 28534525
- PMCID: PMC5457521
- DOI: 10.1038/ncomms15411
Covalently linked dengue virus envelope glycoprotein dimers reduce exposure of the immunodominant fusion loop epitope
Abstract
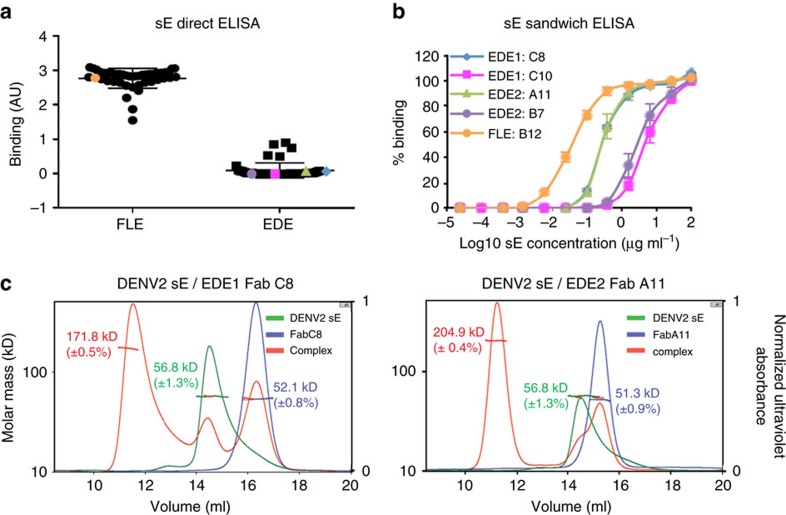
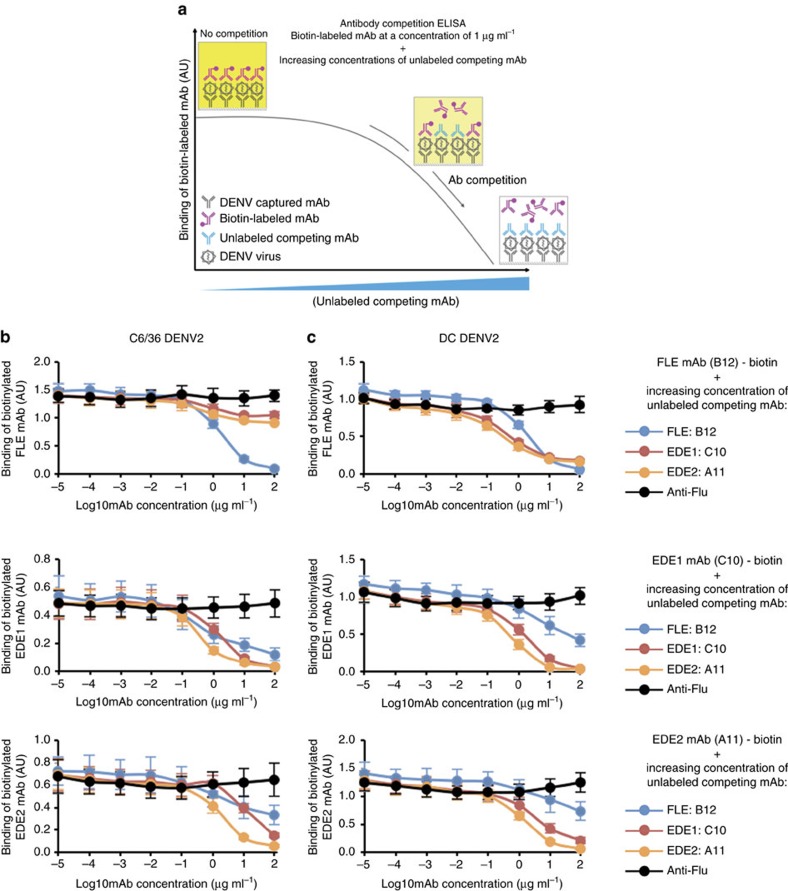
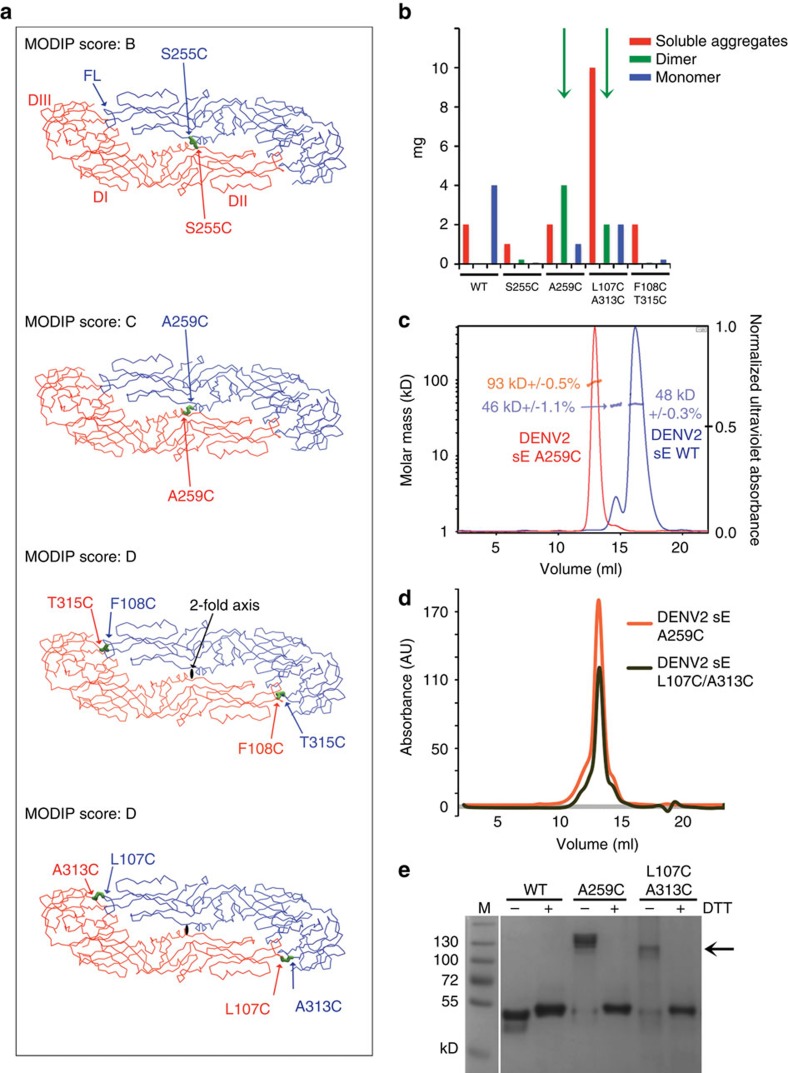
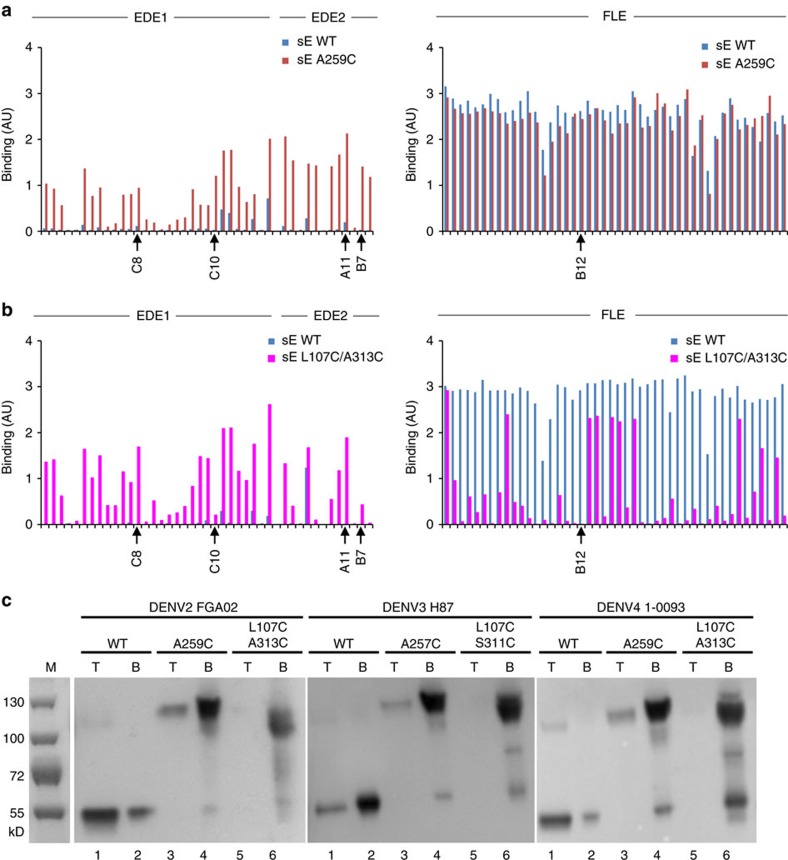
A problem in the search for an efficient vaccine against dengue virus is the immunodominance of the fusion loop epitope (FLE), a segment of the envelope protein E that is buried at the interface of the E dimers coating mature viral particles. Anti-FLE antibodies are broadly cross-reactive but poorly neutralizing, displaying a strong infection enhancing potential. FLE exposure takes place via dynamic 'breathing' of E dimers at the virion surface. In contrast, antibodies targeting the E dimer epitope (EDE), readily exposed at the E dimer interface over the region of the conserved fusion loop, are very potent and broadly neutralizing. We here engineer E dimers locked by inter-subunit disulfide bonds, and show by X-ray crystallography and by binding to a panel of human antibodies that these engineered dimers do not expose the FLE, while retaining the EDE exposure. These locked dimers are strong immunogen candidates for a next-generation vaccine.
Conflict of interest statement
The EDE antibodies, EDE epitope and envelope protein dimers that induce EDE antibodies are the subject of a patent application by Imperial College and Institute Pasteur on which G.S., J.M., F.A.R., A.R., G.B.-S., P.G.-C., M.-C.V. and S.D. are named as inventors. The remaining authors declare no competing financial interests.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Screaton G., Mongkolsapaya J., Yacoub S. & Roberts C. New insights into the immunopathology and control of dengue virus infection. Nat. Rev. Immunol. 15, 745–759 (2015). - PubMed
-
- Guzman M. G., Gubler D. J., Izquierdo A., Martinez E. & Halstead S. B. Dengue infection. Nat. Rev. Dis. Primers 2, 16055 (2016). - PubMed
-
- Simmons C. P., Farrar J. J., Nguyen v. ,V. & Wills B. Dengue. N. Engl. J. Med. 366, 1423–1432 (2012). - PubMed
-
- World Health Organization (WHO). Strategic Advisory Group of Experts. Dengue vaccine http://www.who.int/immunization/sage/meetings/2016/april/SAGE_April_2016... (2016).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources