Zika Virus Persistence in the Central Nervous System and Lymph Nodes of Rhesus Monkeys
- PMID: 28457610
- PMCID: PMC5426912
- DOI: 10.1016/j.cell.201704008
Zika Virus Persistence in the Central Nervous System and Lymph Nodes of Rhesus Monkeys
Abstract
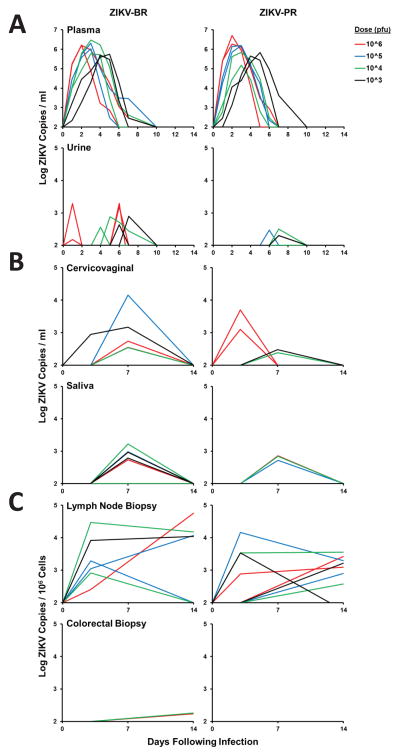
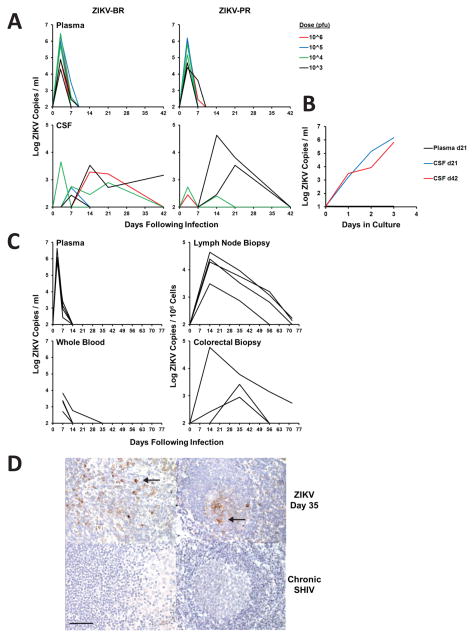
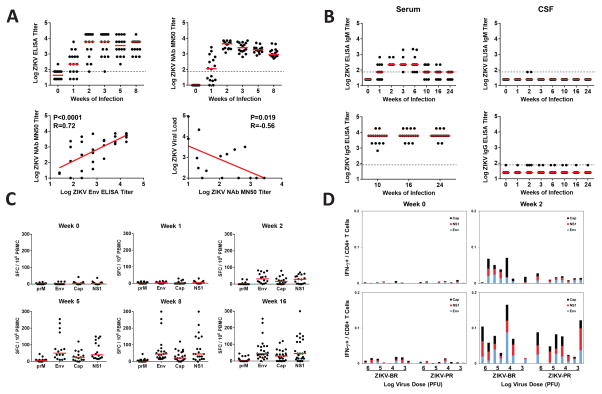
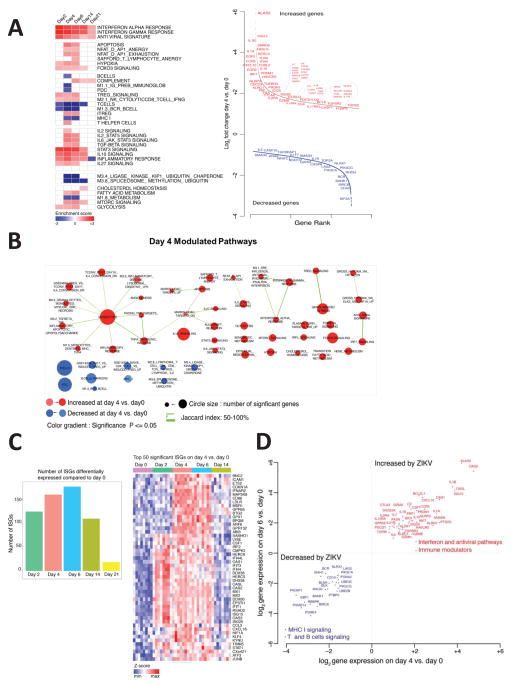
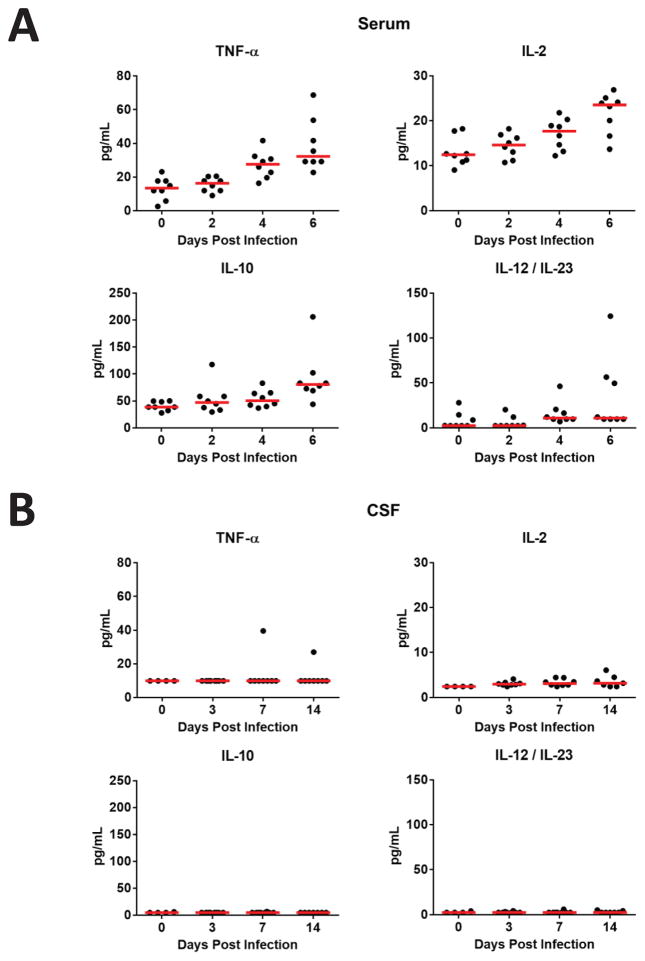
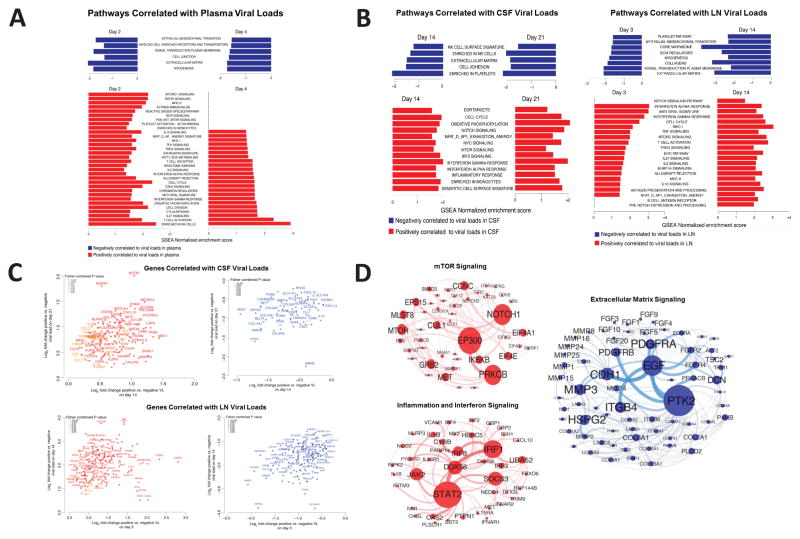
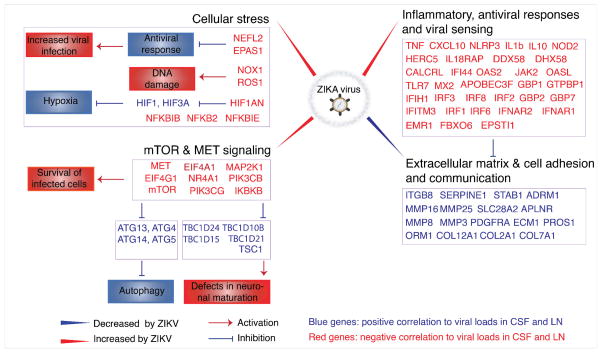
Zika virus (ZIKV) is associated with severe neuropathology in neonates as well as Guillain-Barré syndrome and other neurologic disorders in adults. Prolonged viral shedding has been reported in semen, suggesting the presence of anatomic viral reservoirs. Here we show that ZIKV can persist in cerebrospinal fluid (CSF) and lymph nodes (LN) of infected rhesus monkeys for weeks after virus has been cleared from peripheral blood, urine, and mucosal secretions. ZIKV-specific neutralizing antibodies correlated with rapid clearance of virus in peripheral blood but remained undetectable in CSF for the duration of the study. Viral persistence in both CSF and LN correlated with upregulation of mechanistic target of rapamycin (mTOR), proinflammatory, and anti-apoptotic signaling pathways, as well as downregulation of extracellular matrix signaling pathways. These data raise the possibility that persistent or occult neurologic and lymphoid disease may occur following clearance of peripheral virus in ZIKV-infected individuals.
Keywords: CSF; Zika; infection; lymph node; mTOR; persistence; rhesus; sanctuaries; transcriptomics; viral dynamics.
Copyright © 2017 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Barba-Spaeth G, Dejnirattisai W, Rouvinski A, Vaney MC, Medits I, Sharma A, Simon-Loriere E, Sakuntabhai A, Cao-Lormeau VM, Haouz A, et al. Structural basis of potent Zika-dengue virus antibody cross-neutralization. Nature. 2016;536:48–53. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous