Identification of Zika virus epitopes reveals immunodominant and protective roles for dengue virus cross-reactive CD8+ T cells
- PMID: 28288094
- PMCID: PMC5918137
- DOI: 10.1038/nmicrobiol.2017.36
Identification of Zika virus epitopes reveals immunodominant and protective roles for dengue virus cross-reactive CD8+ T cells
Abstract
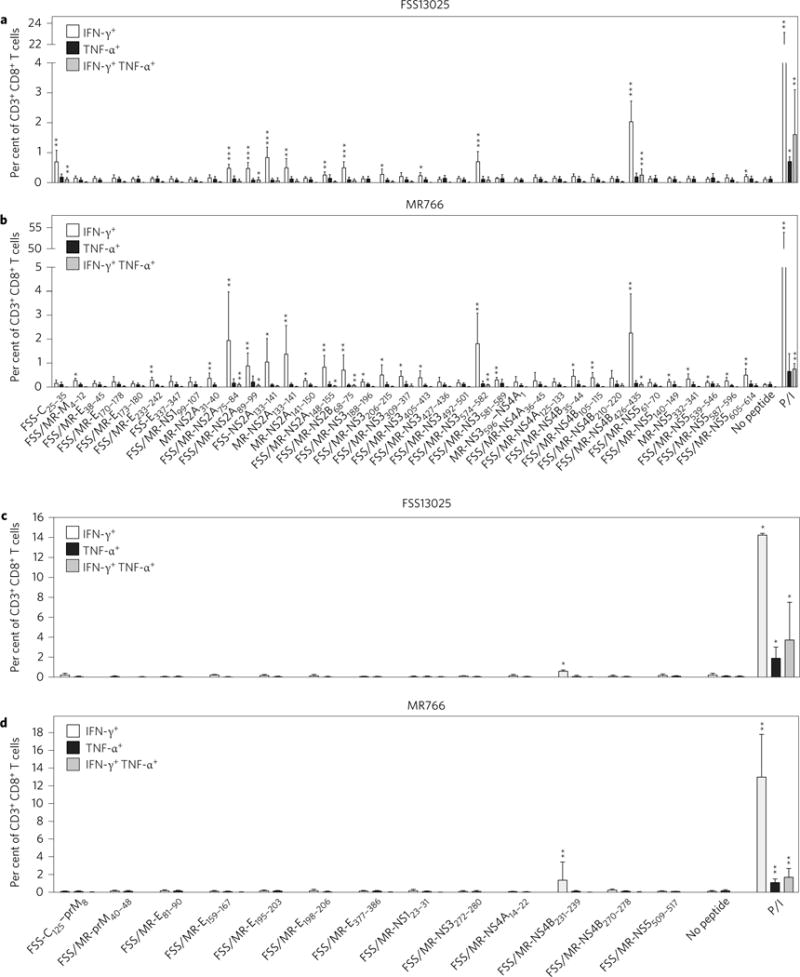
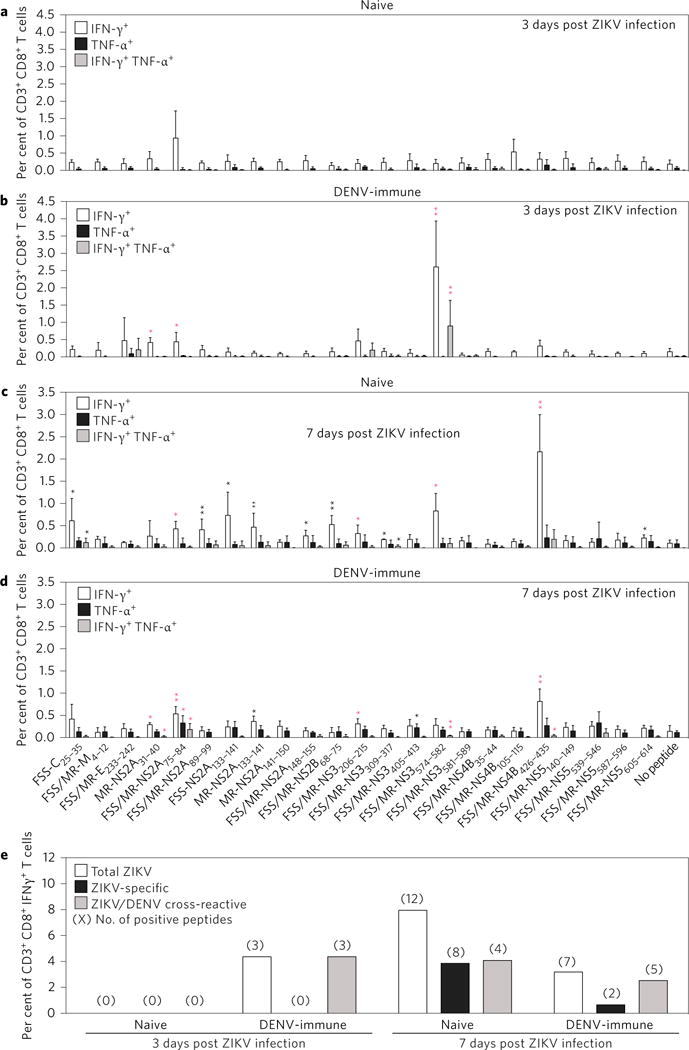
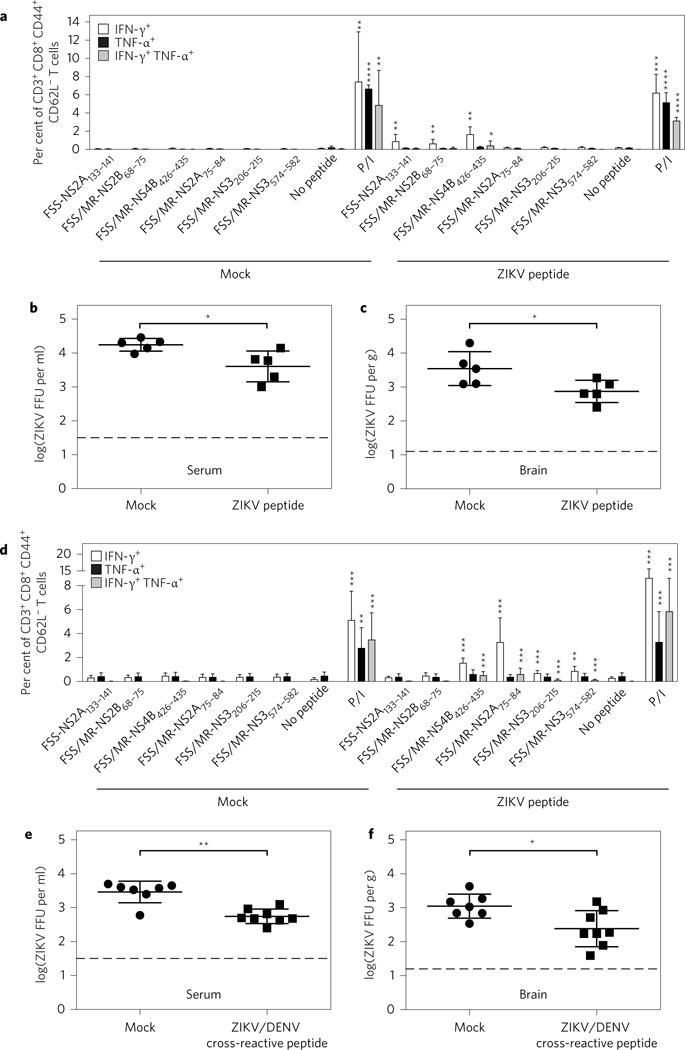
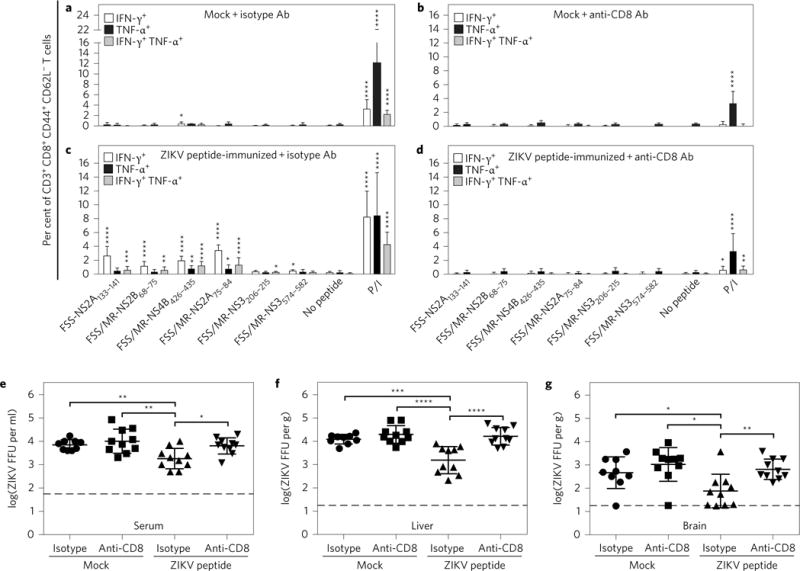
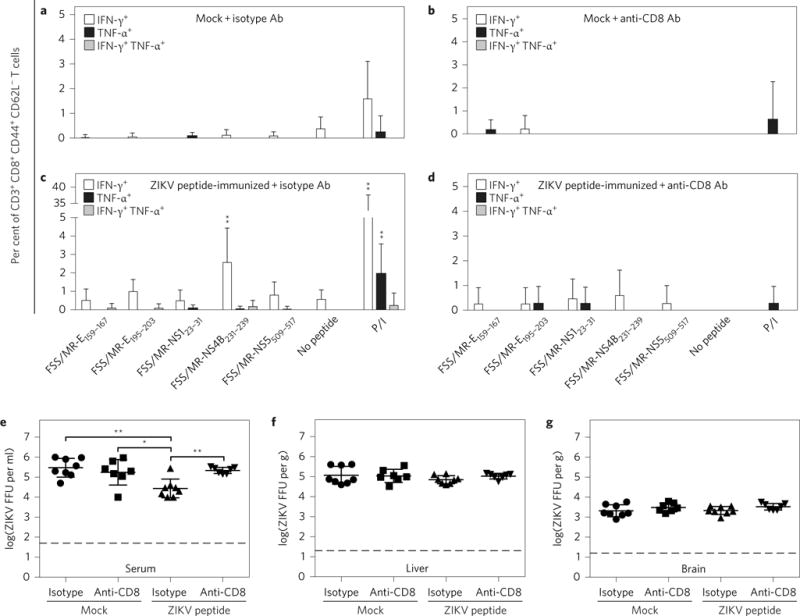
CD8+ T cells play an important role in controlling Flavivirus infection, including Zika virus (ZIKV). Here, we have identified 25 HLA-B*0702-restricted epitopes and 1 HLA-A*0101-restricted epitope using interferon (IFN)-γ enzyme-linked immunospot (ELISPOT) and intracellular cytokine staining (ICS) in ZIKV-infected IFN-α/β receptor-deficient HLA transgenic mice. The cross-reactivity of ZIKV epitopes to dengue virus (DENV) was tested using IFN-γ-ELISPOT and IFN-γ-ICS on CD8+ T cells from DENV-infected mice, and five cross-reactive HLA-B*0702-binding peptides were identified by both assays. ZIKV/DENV cross-reactive CD8+ T cells in DENV-immune mice expanded post ZIKV challenge and dominated in the subsequent CD8+ T cell response. ZIKV challenge following immunization of mice with ZIKV-specific and ZIKV/DENV cross-reactive epitopes elicited CD8+ T cell responses that reduced infectious ZIKV levels, and CD8+ T cell depletions confirmed that CD8+ T cells mediated this protection. These results identify ZIKV-specific and ZIKV/DENV cross-reactive epitopes and demonstrate both an altered immunodominance pattern in the DENV-immune setting relative to naive, as well as a protective role for epitope-specific CD8+ T cells against ZIKV. These results have important implications for ZIKV vaccine development and provide a mouse model for evaluating anti-ZIKV CD8+ T cell responses of human relevance.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Comment in
-
Host response: Cross-fit T cells battle Zika virus.Collins M, de Silva A. Collins M, et al. Nat Microbiol. 2017 May 25;2:17082. doi: 10.1038/nmicrobiol.2017.82. Nat Microbiol. 2017. PMID: 28540930 No abstract available.
References
-
- Choumet V, Despres P. Dengue and other flavivirus infections. Rev Sci Tech. 2015;34:473–478. 467–472. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials