Leishmania (Leishmania) amazonensis induces macrophage miR-294 and miR-721 expression and modulates infection by targeting NOS2 and L-arginine metabolism
- PMID: 28276497
- PMCID: PMC5343489
- DOI: 10.1038/srep44141
Leishmania (Leishmania) amazonensis induces macrophage miR-294 and miR-721 expression and modulates infection by targeting NOS2 and L-arginine metabolism
Abstract
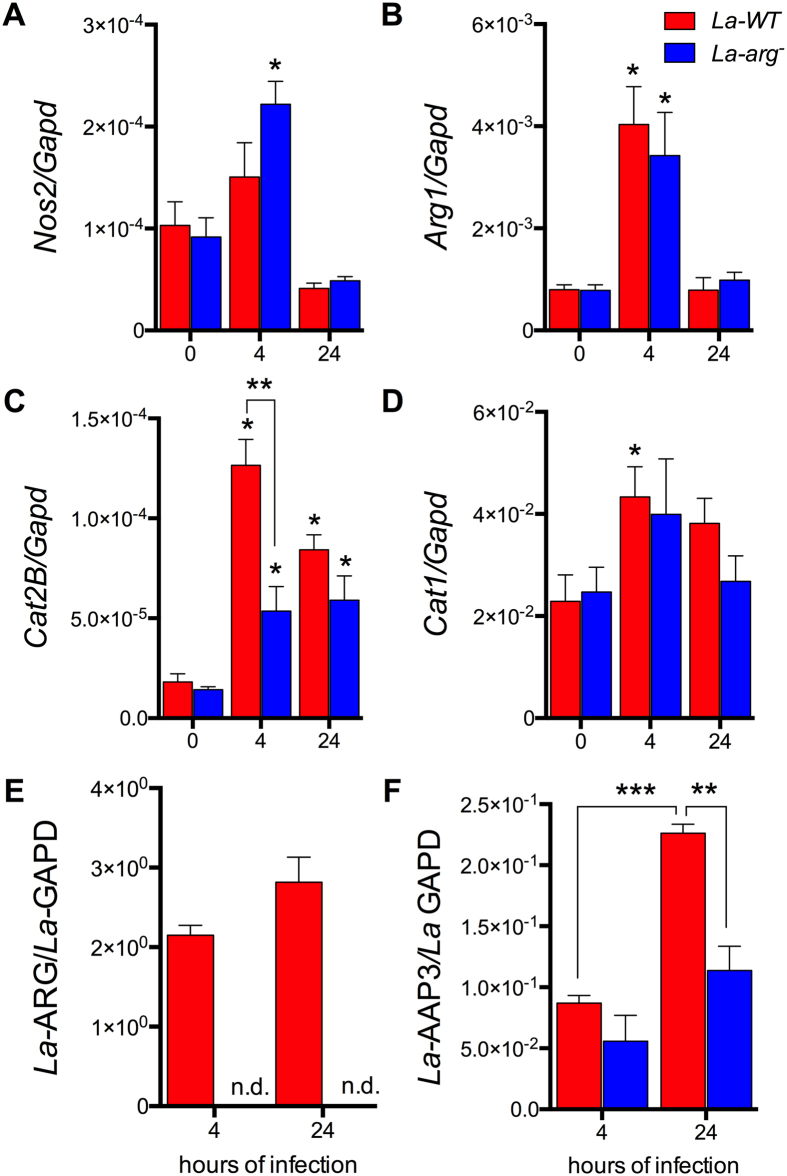
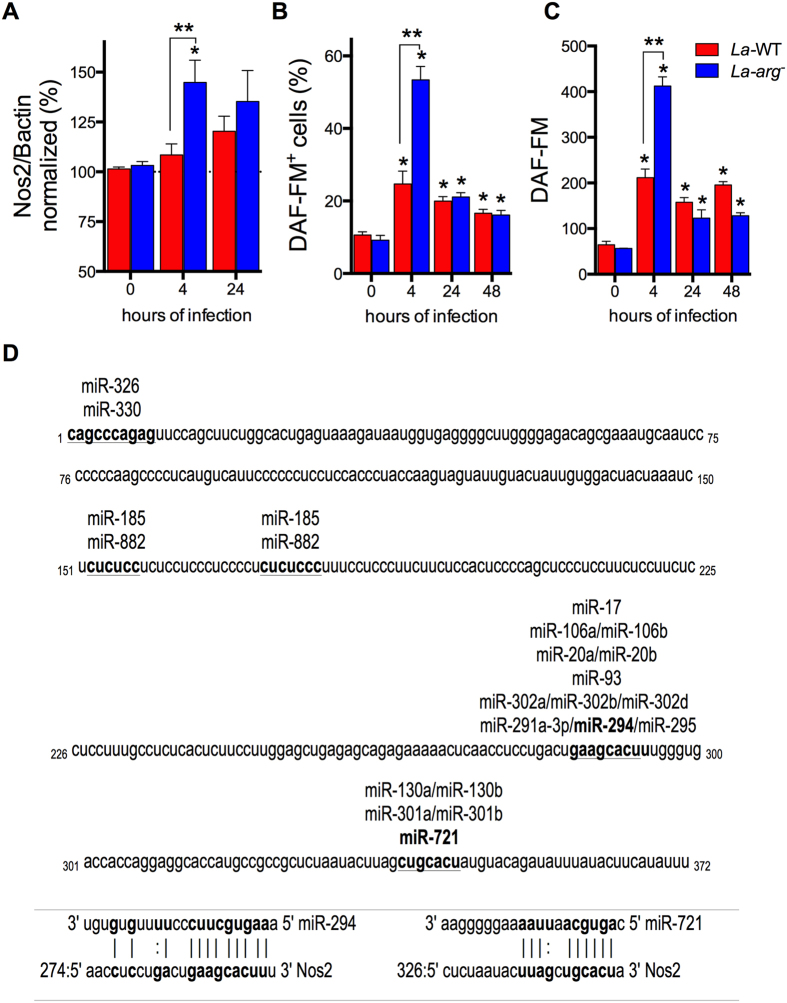
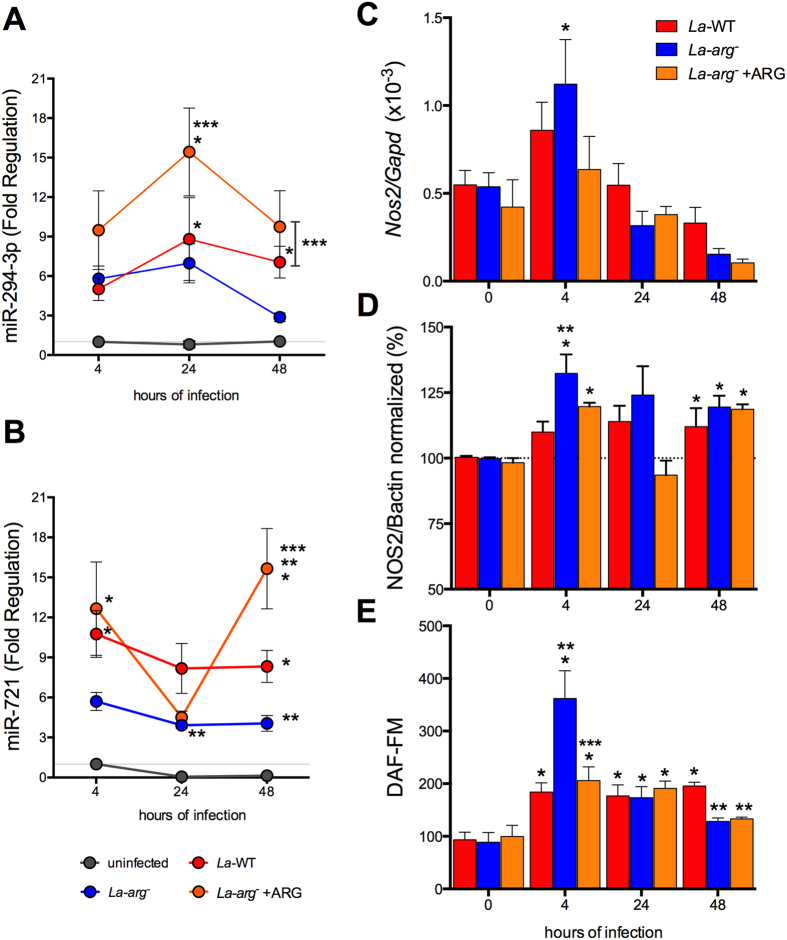
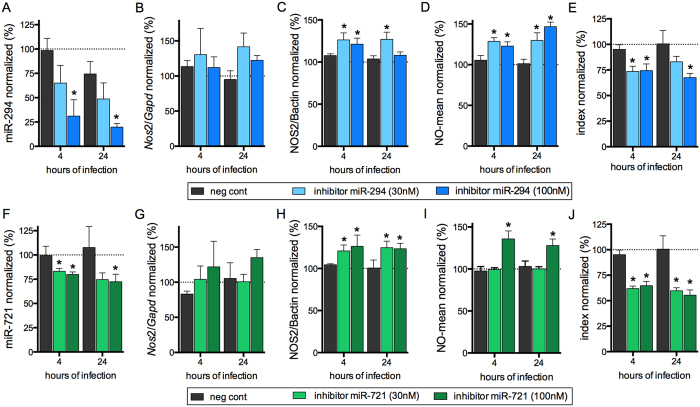
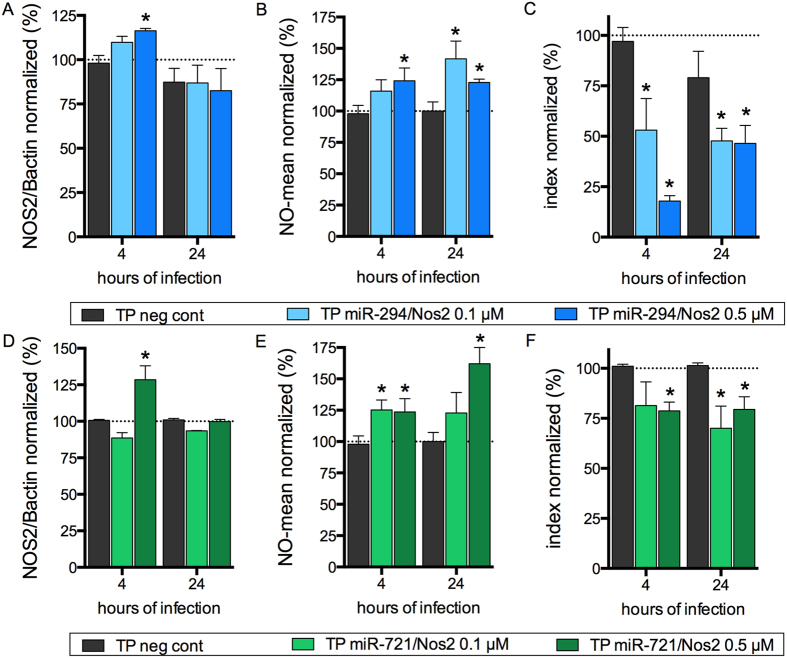
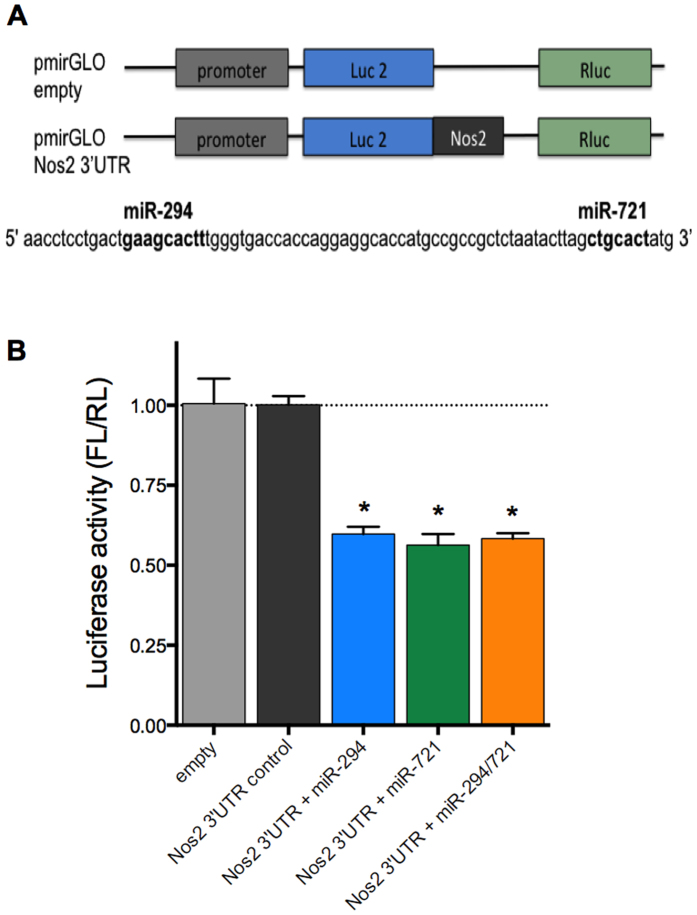
Leishmania (Leishmania) amazonensis is an intracellular protozoan parasite responsible for the cutaneous leishmaniasis. The parasite replicates inside mammalian macrophage to establish infection. Host-pathogen interactions result in microRNA-mediated post-transcriptional regulation of host genes involved in inflammatory immune response. We analyzed macrophage miRNA profiles during L. (L.) amazonensis infection. The regulation of macrophage miRNA expression by the parasite correlates with/depends on parasite arginase activity during infection. L. (L.) amazonensis (La-WT) presented significant miRNA profile alteration (27%) compared to L. (L.) amazonensis arginase knockout (La-arg-) (~40%) in relation to uninfected-macrophages. We observed that 78% of the altered miRNAs were up-regulated in La-WT infection, while only 32% were up-regulated in La-arg--infected macrophages. In contrast to La-WT, the lack of L. (L.) amazonensis arginase led to the inhibition of miR-294 and miR-721 expression. The expression of miR-294 and miR-721 was recovered to levels similar to La-WT in La-arg- addback mutant. The inhibition of miR-294/Nos2 and miR721/Nos2 interactions increased NOS2 expression and NO production, and reduced L. (L.) amazonensis infectivity, confirming Nos2 as target of these miRNAs. The role of miR-294 and miR-721 in the regulation of NOS2 expression during Leishmania replication in infected macrophages pointing these miRNAs as potential new targets for drug development.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Green S. J., Crawford R. M., Hockmeyer J. T., Meltzer M. S. & Nacy C. A. Leishmania major amastigotes initiate the L-arginine-dependent killing mechanism in IFN-gamma-stimulated macrophages by induction of tumor necrosis factor-alpha. J Immunol 145, 4290–4297 (1990). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical