Exosome secretion affects social motility in Trypanosoma brucei
- PMID: 28257521
- PMCID: PMC5352147
- DOI: 10.1371/journal.ppat.1006245
Exosome secretion affects social motility in Trypanosoma brucei
Abstract
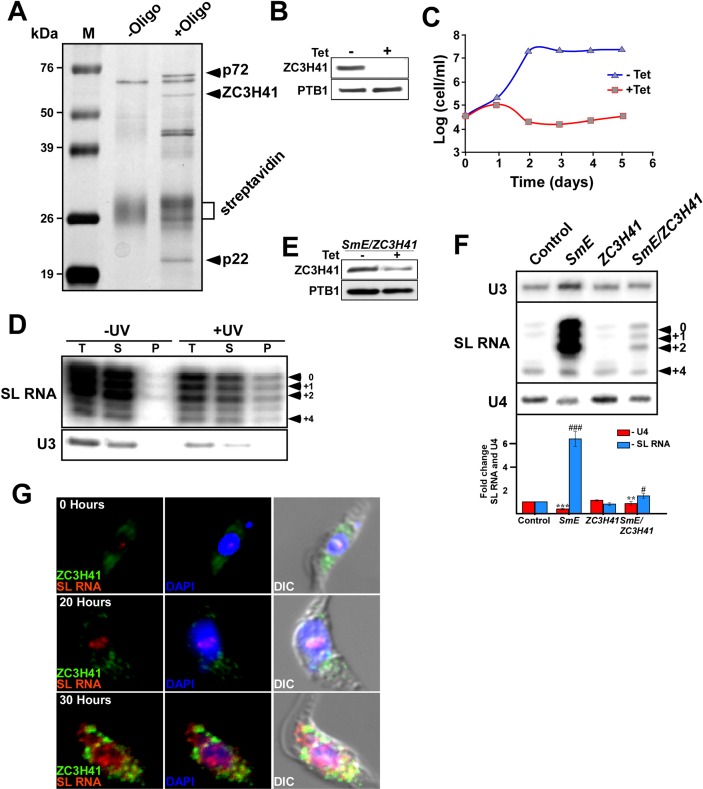
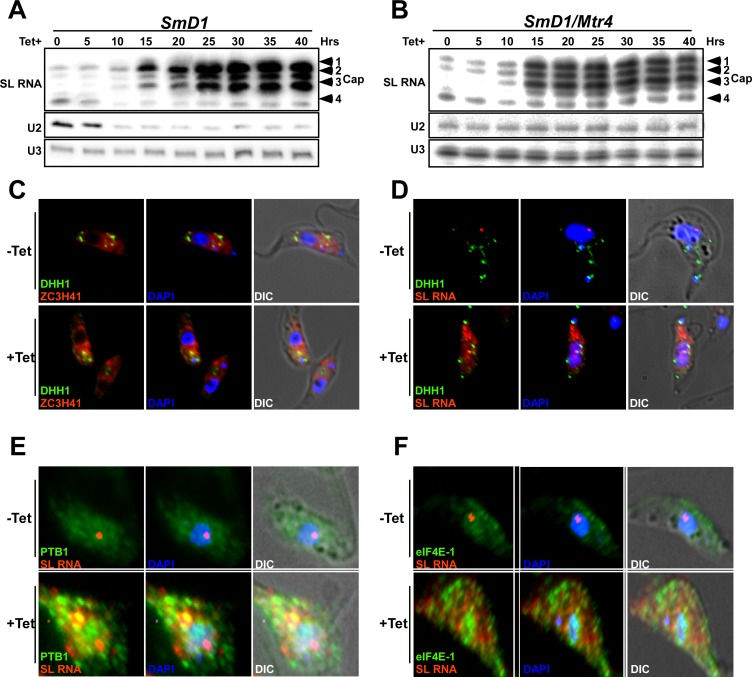
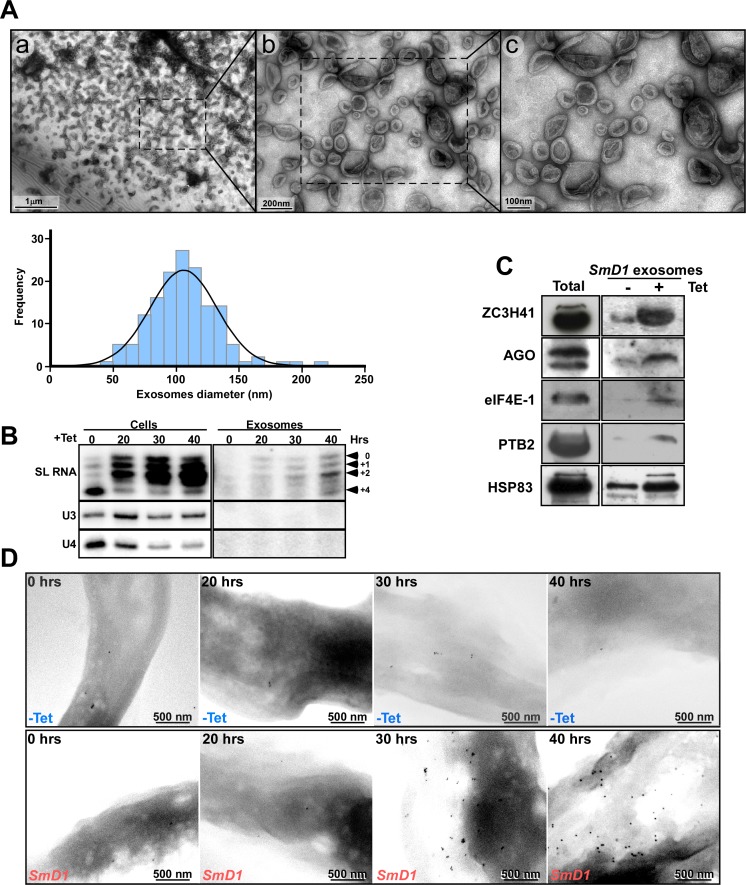
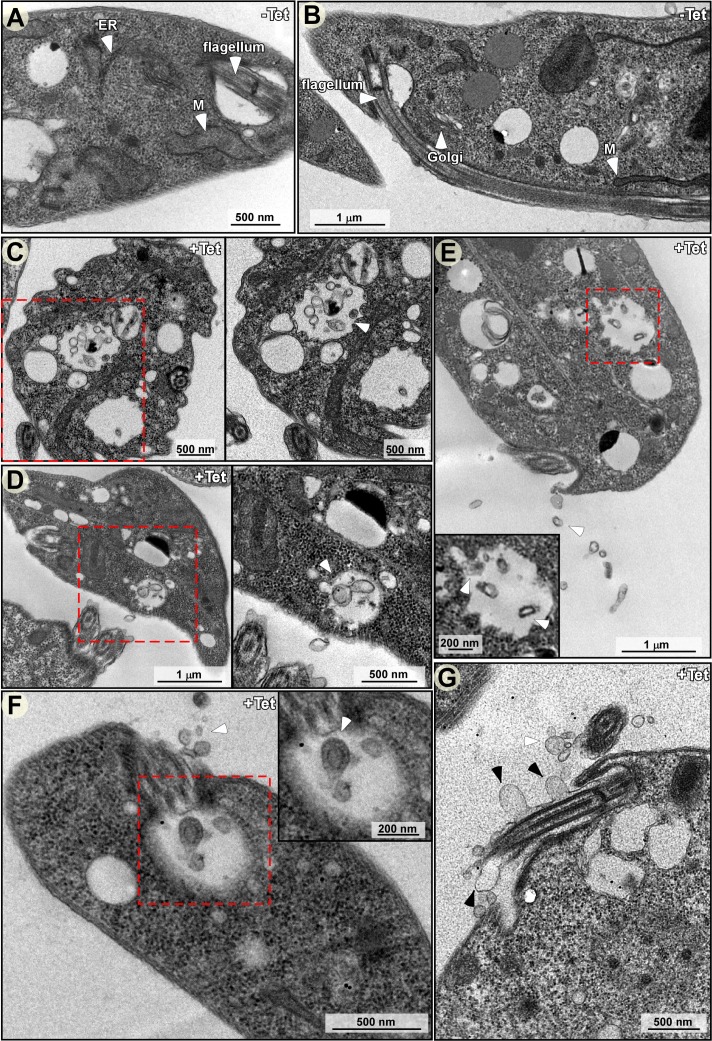
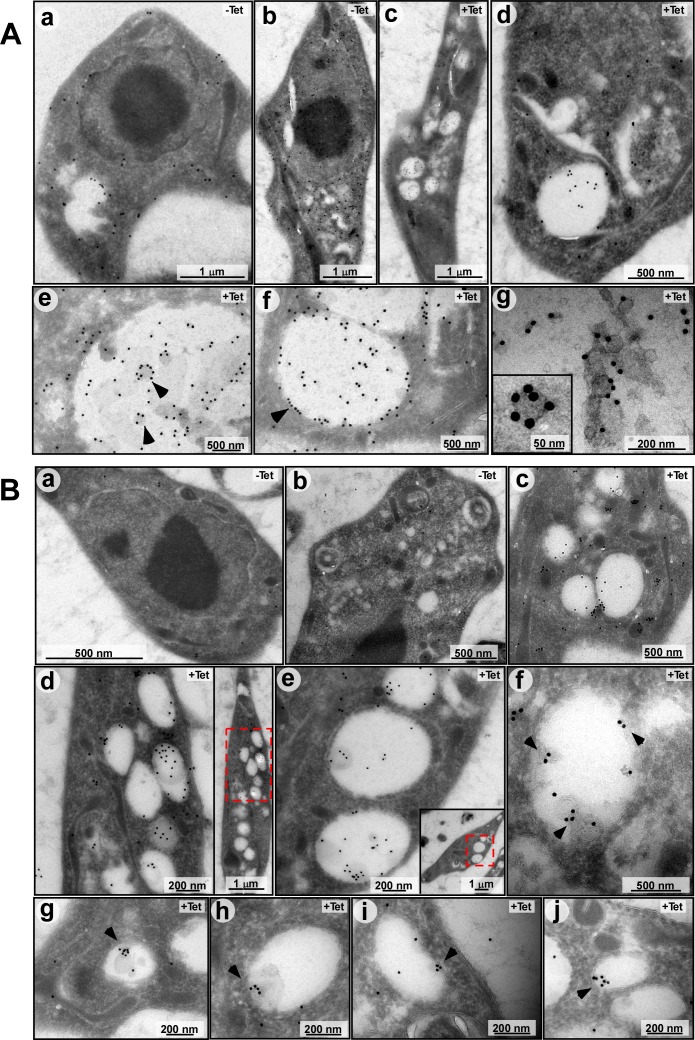
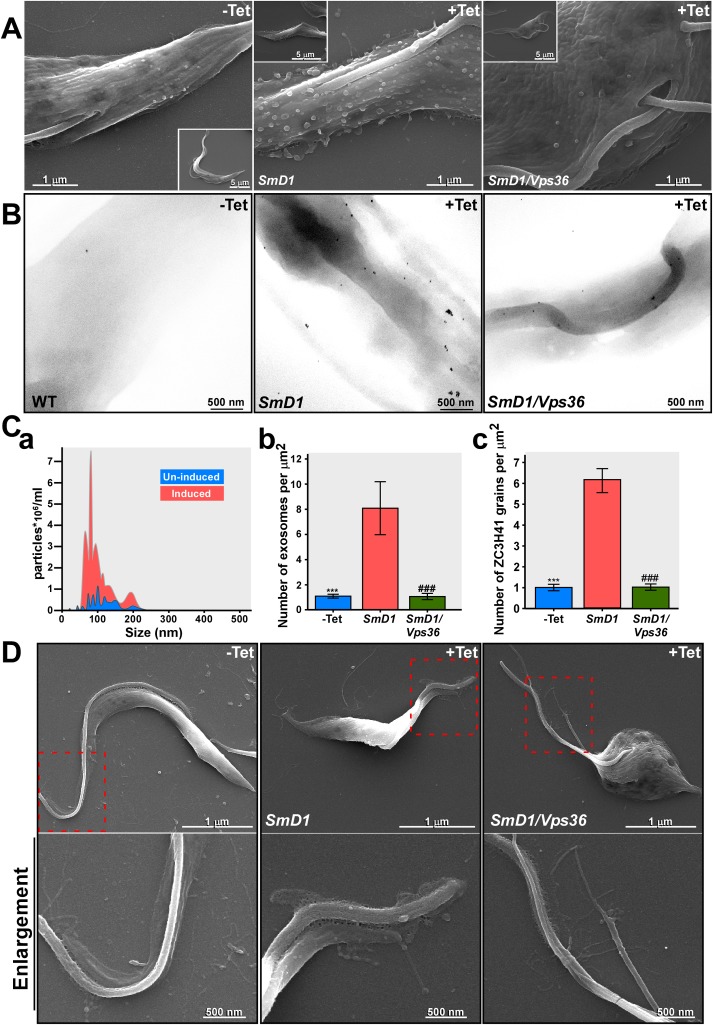
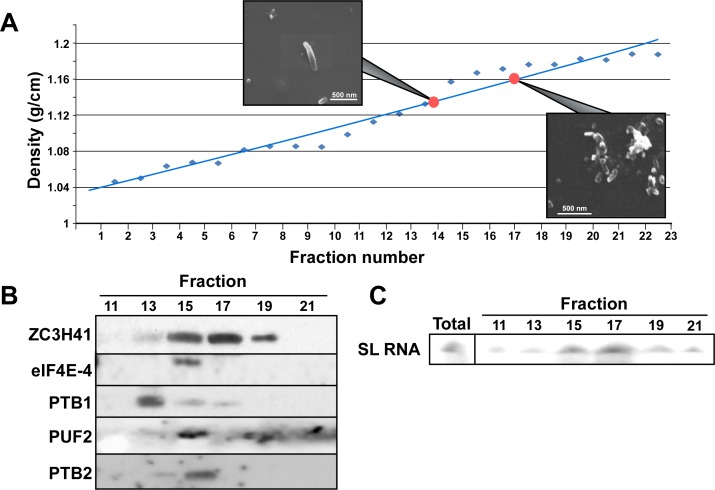
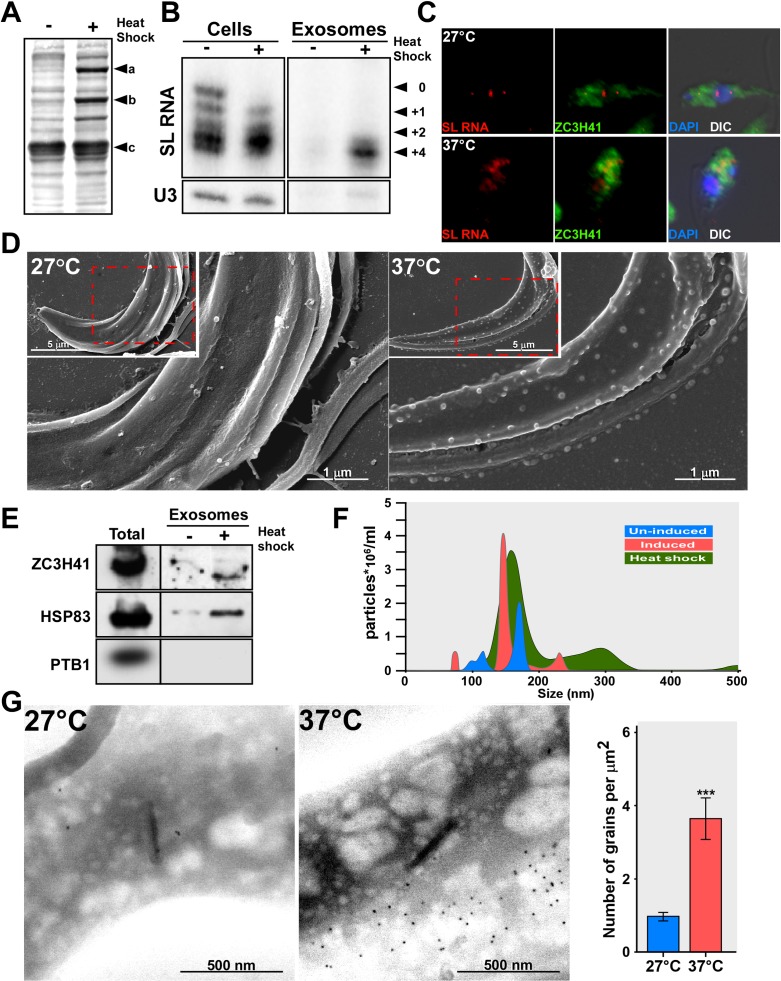
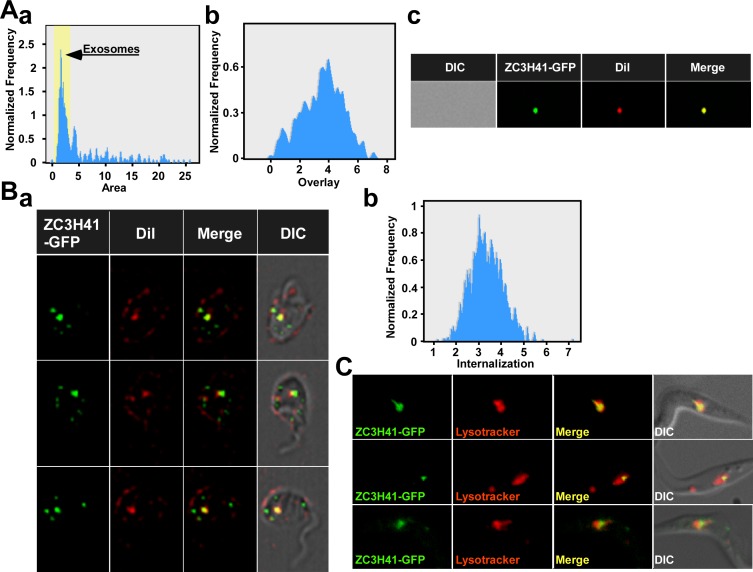
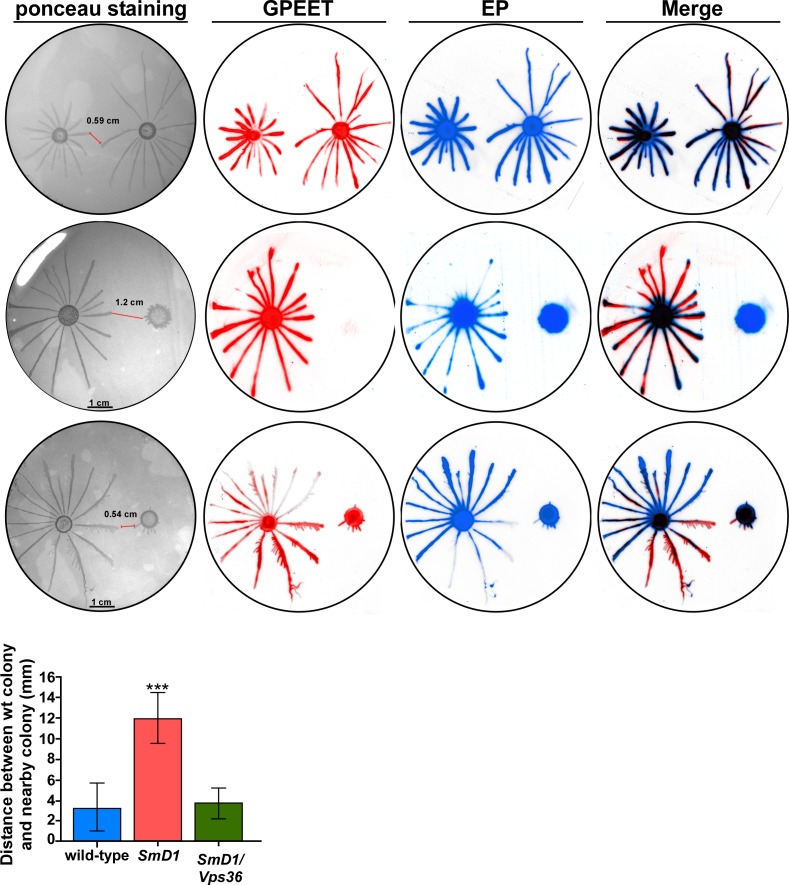
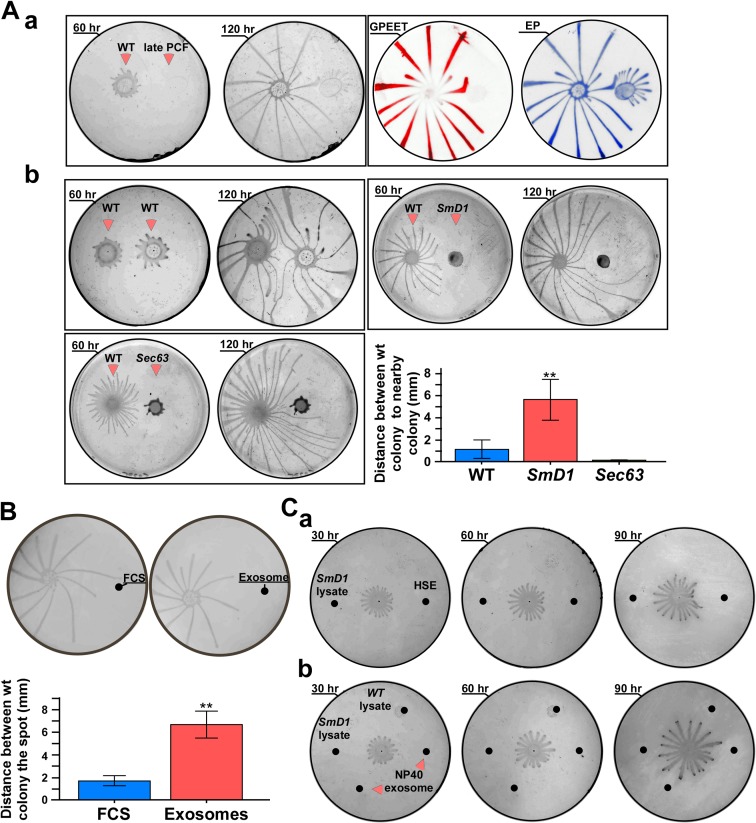
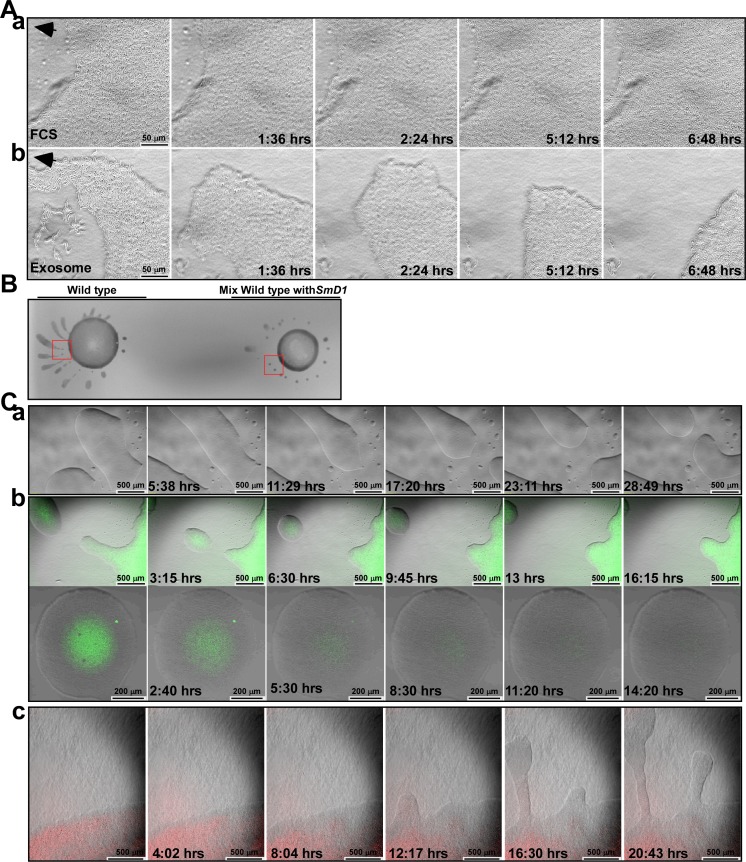
Extracellular vesicles (EV) secreted by pathogens function in a variety of biological processes. Here, we demonstrate that in the protozoan parasite Trypanosoma brucei, exosome secretion is induced by stress that affects trans-splicing. Following perturbations in biogenesis of spliced leader RNA, which donates its spliced leader (SL) exon to all mRNAs, or after heat-shock, the SL RNA is exported to the cytoplasm and forms distinct granules, which are then secreted by exosomes. The exosomes are formed in multivesicular bodies (MVB) utilizing the endosomal sorting complexes required for transport (ESCRT), through a mechanism similar to microRNA secretion in mammalian cells. Silencing of the ESCRT factor, Vps36, compromised exosome secretion but not the secretion of vesicles derived from nanotubes. The exosomes enter recipient trypanosome cells. Time-lapse microscopy demonstrated that cells secreting exosomes or purified intact exosomes affect social motility (SoMo). This study demonstrates that exosomes are delivered to trypanosome cells and can change their migration. Exosomes are used to transmit stress signals for communication between parasites.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Van Den Abbeele J, Claes Y, van Bockstaele D, Le Ray D, Coosemans M. Trypanosoma brucei spp. development in the tsetse fly: characterization of the post-mesocyclic stages in the foregut and proboscis. Parasitology. 1999;118:469–78. Epub 1999年06月11日. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources