Levels of insecticide resistance to deltamethrin, malathion, and temephos, and associated mechanisms in Aedes aegypti mosquitoes from the Guadeloupe and Saint Martin islands (French West Indies)
- PMID: 28187780
- PMCID: PMC5303256
- DOI: 10.1186/s40249-017-0254-x
Levels of insecticide resistance to deltamethrin, malathion, and temephos, and associated mechanisms in Aedes aegypti mosquitoes from the Guadeloupe and Saint Martin islands (French West Indies)
Abstract
Background: In the Guadeloupe and Saint Martin islands, Aedes aegypti mosquitoes are the only recognized vectors of dengue, chikungunya, and Zika viruses. For around 40 years, malathion was used as a mosquito adulticide and temephos as a larvicide. Since the European Union banned the use of these two insecticide molecules in the first decade of the 21st century, deltamethrin and Bacillus thuringiensis var. israelensis are the remaining adulticide and larvicide, respectively, used in Guadeloupe. In order to improve the management of vector control activities in Guadeloupe and Saint Martin, we investigated Ae. aegypti resistance to and mechanisms associated with deltamethrin, malathion, and temephos.
Methods: Ae. aegypti mosquitoes were collected from six different localities of Guadeloupe and Saint Martin. Larvae were used for malathion and temephos bioassays, and adult mosquitoes for deltamethrin bioassays, following World Health Organization recommendations. Knockdown resistance (Kdr) genotyping for V1016I and F1534C mutations, and expression levels of eight enzymes involved in detoxification mechanisms were examined in comparison with the susceptible reference Bora Bora strain.
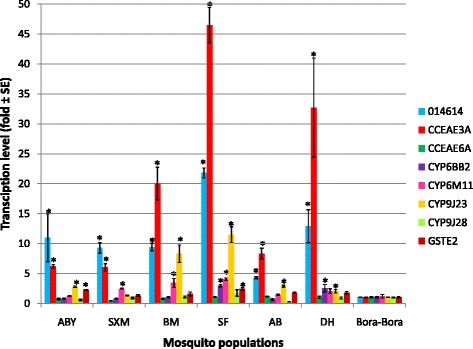
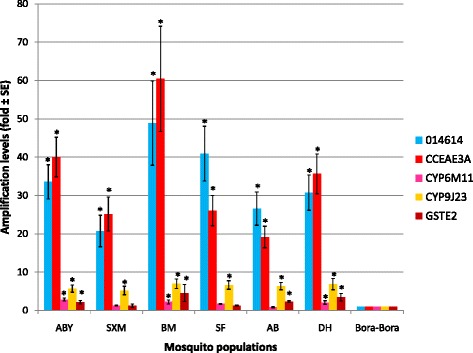
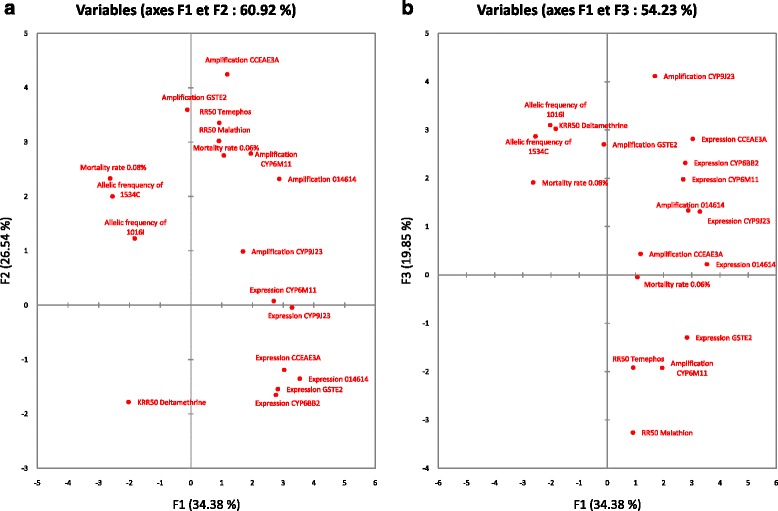
Results: Resistance ratios (RR50) calculated for Ae. aegypti larvae showed high resistance levels to temephos (from 8.9 to 33.1-fold) and low resistance levels to malathion (from 1.7 to 4.4-fold). Adult females displayed moderate resistance levels to deltamethrin regarding the time necessary to affect 50% of individuals, varying from 8.0 to 28.1-fold. Molecular investigations on adult mosquitoes showed high resistant allele frequencies for V1016I and F1534C (from 85 to 96% and from 90 to 98%, respectively), as well as an overexpression of the glutathione S-transferase gene, GSTe2, the carboxylesterase CCEae3a, and the cytochrome genes 014614, CYP6BB2, CYP6M11, and CYP9J23.
Conclusions: Ae. aegypti populations from Guadeloupe and Saint Martin exhibit multiple resistance to organophosphates (temephos and malathion), and pyrethroids (deltamethrin). The mechanisms associated with these resistance patterns show strong frequencies of F1534C and V1016I Kdr mutations, and an over-expression of CCEae3a, GSTe2, and four cytochrome P450 genes (014614, CYP9J23, CYP6M11, CYP6BB2). These results will form the baseline for a deeper understanding of the insecticide resistance levels and associated mechanisms of Ae. aegypti populations and will be used to improve vector control strategies in Guadeloupe and Saint Martin.
Keywords: Aedes aegypti; Deltamethrin; Guadeloupe; Insecticide resistance; Malathion; Mosquitoes; Saint Martin; Temephos.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- CIRE ANTILLES-GUYANE. Emergence du virus Zika aux Antilles Guyane. Situation épidémiologique. INVS, Point épidémiologique du 28 avril 2016: 16/2016.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases