Type Three Secretion System in Attaching and Effacing Pathogens
- PMID: 27818950
- PMCID: PMC5073101
- DOI: 10.3389/fcimb.2016.00129
Type Three Secretion System in Attaching and Effacing Pathogens
Abstract
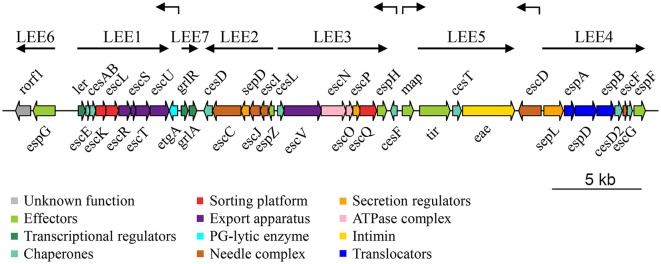
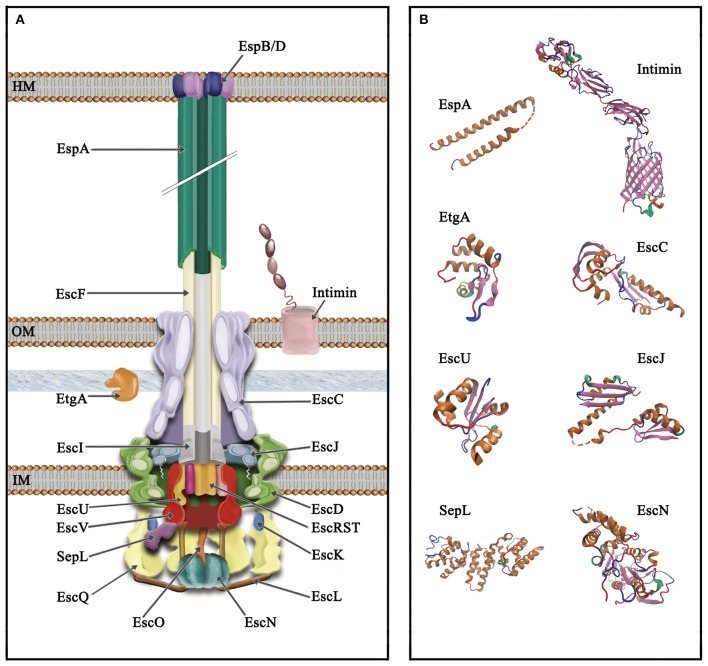
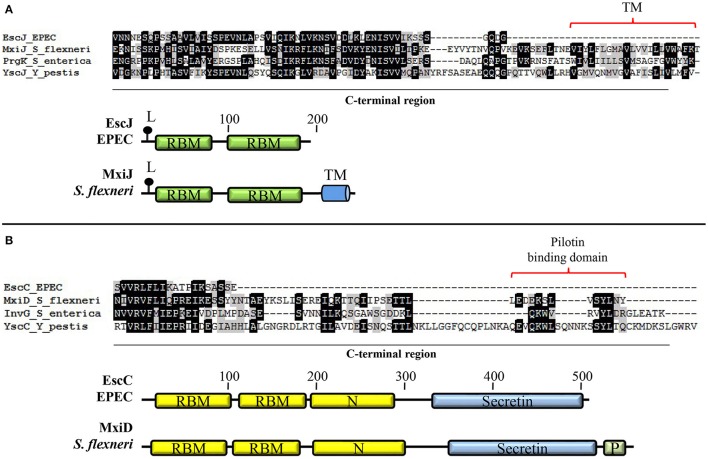
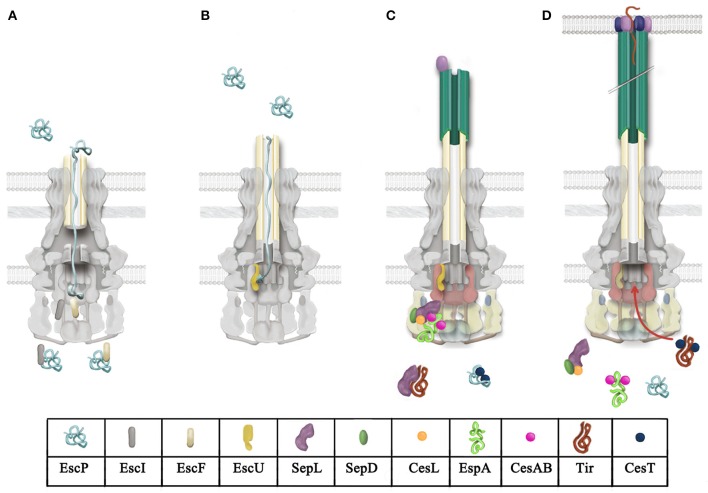
Enteropathogenic Escherichia coli and enterohemorrhagic E. coli are diarrheagenic bacterial human pathogens that cause severe gastroenteritis. These enteric pathotypes, together with the mouse pathogen Citrobacter rodentium, belong to the family of attaching and effacing pathogens that form a distinctive histological lesion in the intestinal epithelium. The virulence of these bacteria depends on a type III secretion system (T3SS), which mediates the translocation of effector proteins from the bacterial cytosol into the infected cells. The core architecture of the T3SS consists of a multi-ring basal body embedded in the bacterial membranes, a periplasmic inner rod, a transmembrane export apparatus in the inner membrane, and cytosolic components including an ATPase complex and the C-ring. In addition, two distinct hollow appendages are assembled on the extracellular face of the basal body creating a channel for protein secretion: an approximately 23 nm needle, and a filament that extends up to 600 nm. This filamentous structure allows these pathogens to get through the host cells mucus barrier. Upon contact with the target cell, a translocation pore is assembled in the host membrane through which the effector proteins are injected. Assembly of the T3SS is strictly regulated to ensure proper timing of substrate secretion. The different type III substrates coexist in the bacterial cytoplasm, and their hierarchical secretion is determined by specialized chaperones in coordination with two molecular switches and the so-called sorting platform. In this review, we present recent advances in the understanding of the T3SS in attaching and effacing pathogens.
Keywords: A/E pathogens; Citrobacter rodentium; EHEC; EPEC; injectisome; locus of enterocyte effacement; secretion hierarchy; type III secretion system.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Abe A., de Grado M., Pfuetzner R. A., Sánchez-Sanmartín C., Devinney R., Puente J. L., et al. . (1999). Enteropathogenic Escherichia coli translocated intimin receptor, Tir, requires a specific chaperone for stable secretion. Mol. Microbiol. 33, 1162–1175. - PubMed
-
- Agrain C., Sorg I., Paroz C., Cornelis G. R. (2005). Secretion of YscP from Yersinia enterocolitica is essential to control the length of the injectisome needle but not to change the type III secretion substrate specificity. Mol. Microbiol. 57, 1415–1427. 10.1111/j.1365-2958.2005.04758.x - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources