Climatic and evolutionary drivers of phase shifts in the plague epidemics of colonial India
- PMID: 27791071
- PMCID: PMC5187705
- DOI: 10.1073/pnas.1604985113
Climatic and evolutionary drivers of phase shifts in the plague epidemics of colonial India
Abstract
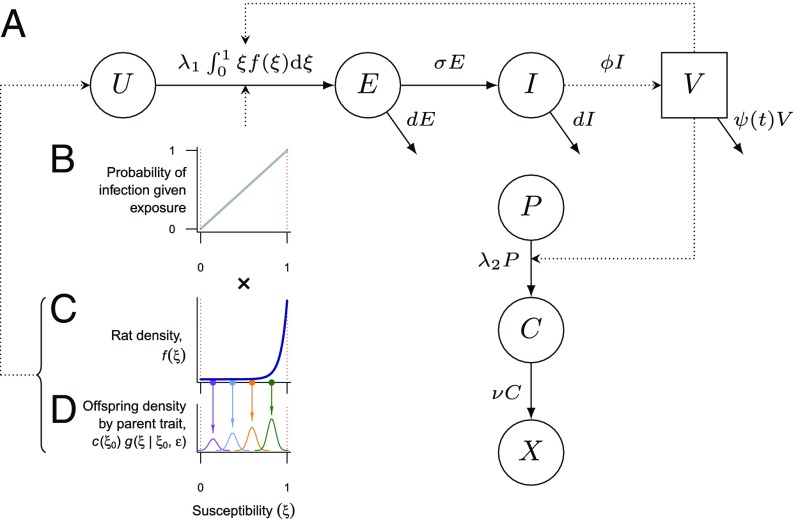
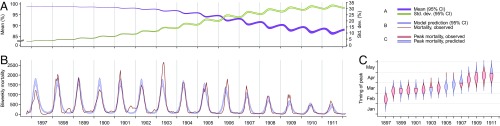
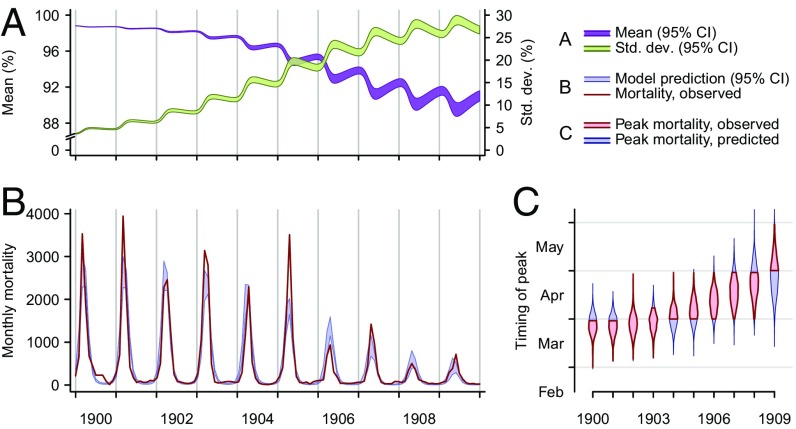
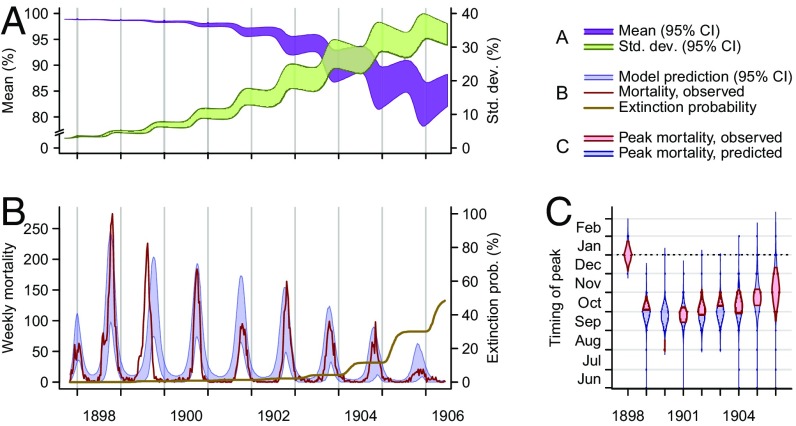
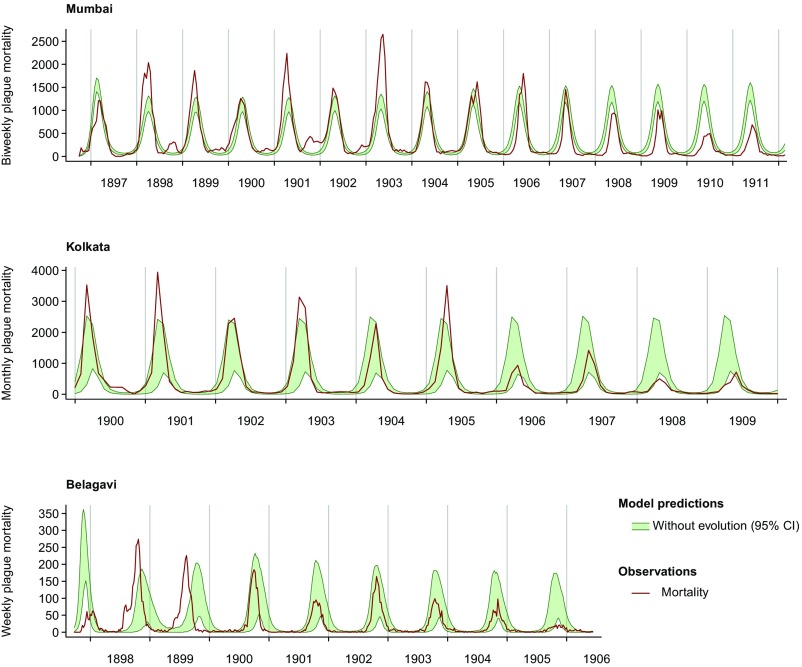
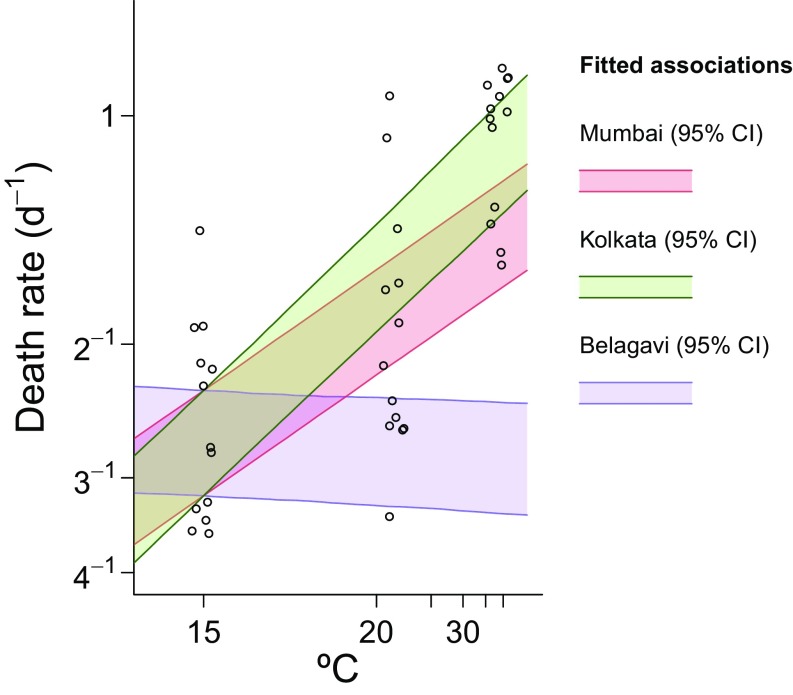
Immune heterogeneity in wild host populations indicates that disease-mediated selection is common in nature. However, the underlying dynamic feedbacks involving the ecology of disease transmission, evolutionary processes, and their interaction with environmental drivers have proven challenging to characterize. Plague presents an optimal system for interrogating such couplings: Yersinia pestis transmission exerts intense selective pressure driving the local persistence of disease resistance among its wildlife hosts in endemic areas. Investigations undertaken in colonial India after the introduction of plague in 1896 suggest that, only a decade after plague arrived, a heritable, plague-resistant phenotype had become prevalent among commensal rats of cities undergoing severe plague epidemics. To understand the possible evolutionary basis of these observations, we developed a mathematical model coupling environmentally forced plague dynamics with evolutionary selection of rats, capitalizing on extensive archival data from Indian Plague Commission investigations. Incorporating increased plague resistance among rats as a consequence of intense natural selection permits the model to reproduce observed changes in seasonal epidemic patterns in several cities and capture experimentally observed associations between climate and flea population dynamics in India. Our model results substantiate Victorian era claims of host evolution based on experimental observations of plague resistance and reveal the buffering effect of such evolution against environmental drivers of transmission. Our analysis shows that historical datasets can yield powerful insights into the transmission dynamics of reemerging disease agents with which we have limited contemporary experience to guide quantitative modeling and inference.
Keywords: immunoecology; infectious disease; modeling; vector-borne disease; zoonosis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Wolinska J, King KC. Environment can alter selection in host-parasite interactions. Trends Parasitol. 2009;25(5):236–244. - PubMed
-
- Sheldon BC, Verhulst S. Ecological immunology: Costly parasite defences and trade-offs in evolutionary ecology. Trends Ecol Evol. 1996;11(8):317–321. - PubMed
-
- Lochmiller RL, Deerenberg C. Trade-offs in evolutionary immunology: Just what is the cost of immunity? Oikos. 2000;88(1):87–98.
-
- Altizer S, Harvell D, Friedle E. Rapid evolutionary dynamics and disease threats to biodiversity. Trends Ecol Evol. 2003;18(11):589–596.
-
- Altizer S, Ostfeld RS, Johnson PTJ, Kutz S, Harvell CD. Climate change and infectious diseases: From evidence to a predictive framework. Science. 2013;341(6145):514–519. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical