Systematic functional analysis of kinases in the fungal pathogen Cryptococcus neoformans
- PMID: 27677328
- PMCID: PMC5052723
- DOI: 10.1038/ncomms12766
Systematic functional analysis of kinases in the fungal pathogen Cryptococcus neoformans
Abstract
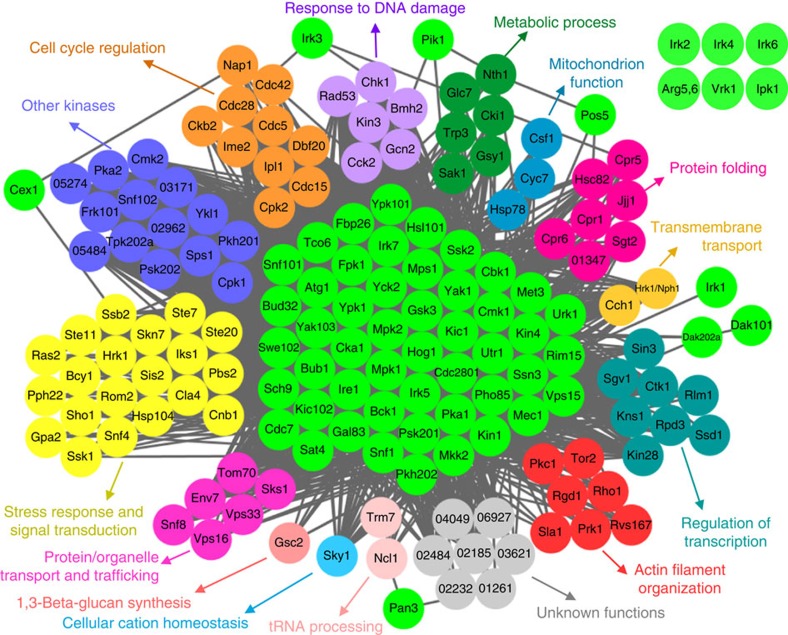
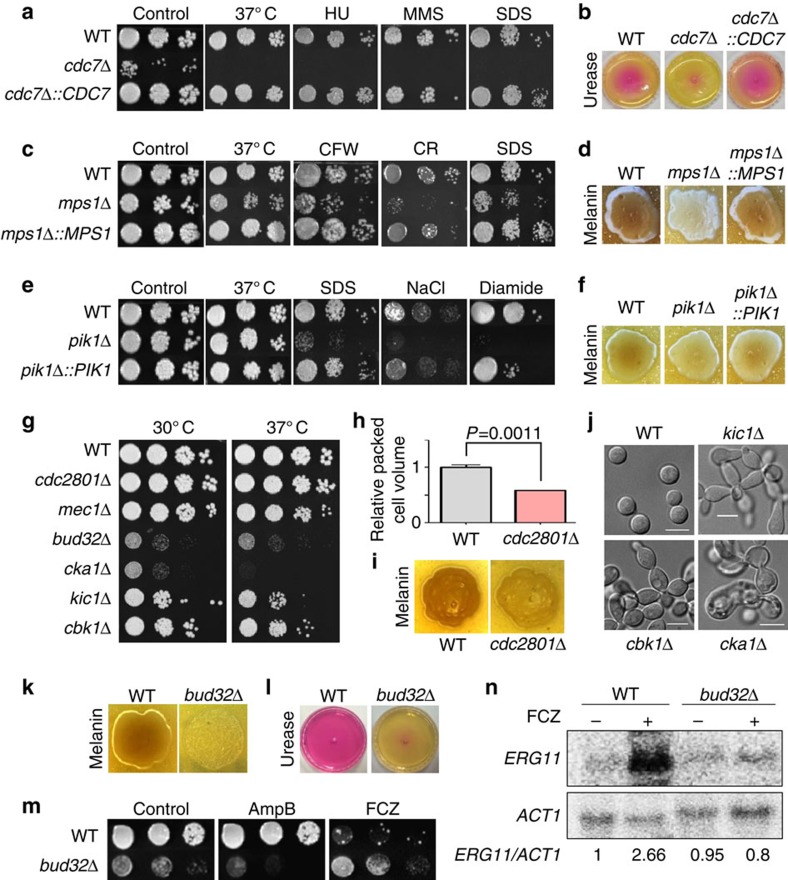
Cryptococcus neoformans is the leading cause of death by fungal meningoencephalitis; however, treatment options remain limited. Here we report the construction of 264 signature-tagged gene-deletion strains for 129 putative kinases, and examine their phenotypic traits under 30 distinct in vitro growth conditions and in two different hosts (insect larvae and mice). Clustering analysis of in vitro phenotypic traits indicates that several of these kinases have roles in known signalling pathways, and identifies hitherto uncharacterized signalling cascades. Virulence assays in the insect and mouse models provide evidence of pathogenicity-related roles for 63 kinases involved in the following biological categories: growth and cell cycle, nutrient metabolism, stress response and adaptation, cell signalling, cell polarity and morphology, vacuole trafficking, transfer RNA (tRNA) modification and other functions. Our study provides insights into the pathobiological signalling circuitry of C. neoformans and identifies potential anticryptococcal or antifungal drug targets.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Park B. J. et al. Estimation of the current global burden of cryptococcal meningitis among persons living with HIV/AIDS. AIDS 23, 525–530 (2009). - PubMed
-
- Lin X. & Heitman J. The biology of the Cryptococcus neoformans species complex. Annu. Rev. Microbiol. 60, 69–105 (2006). - PubMed
-
- Idnurm A. et al. Deciphering the model pathogenic fungus Cryptococcus neoformans. Nat. Rev. Microbiol. 3, 753–764 (2005). - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources