Strategies Used by Bacteria to Grow in Macrophages
- PMID: 27337444
- PMCID: PMC4922531
- DOI: 10.1128/microbiolspec.MCHD-0012-2015
Strategies Used by Bacteria to Grow in Macrophages
Abstract
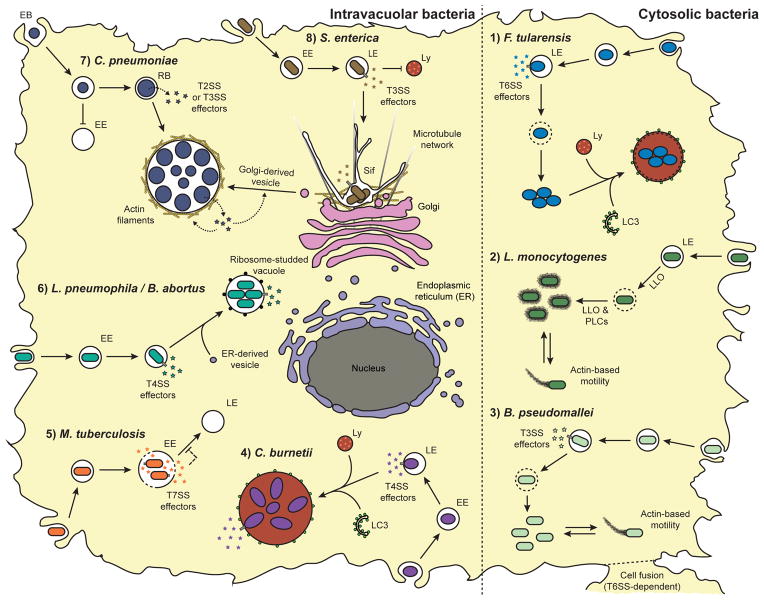
Intracellular bacteria are often clinically relevant pathogens that infect virtually every cell type found in host organisms. However, myeloid cells, especially macrophages, constitute the primary cells targeted by most species of intracellular bacteria. Paradoxically, macrophages possess an extensive antimicrobial arsenal and are efficient at killing microbes. In addition to their ability to detect and signal the presence of pathogens, macrophages sequester and digest microorganisms using the phagolysosomal and autophagy pathways or, ultimately, eliminate themselves through the induction of programmed cell death. Consequently, intracellular bacteria influence numerous host processes and deploy sophisticated strategies to replicate within these host cells. Although most intracellular bacteria have a unique intracellular life cycle, these pathogens are broadly categorized into intravacuolar and cytosolic bacteria. Following phagocytosis, intravacuolar bacteria reside in the host endomembrane system and, to some extent, are protected from the host cytosolic innate immune defenses. However, the intravacuolar lifestyle requires the generation and maintenance of unique specialized bacteria-containing vacuoles and involves a complex network of host-pathogen interactions. Conversely, cytosolic bacteria escape the phagolysosomal pathway and thrive in the nutrient-rich cytosol despite the presence of host cell-autonomous defenses. The understanding of host-pathogen interactions involved in the pathogenesis of intracellular bacteria will continue to provide mechanistic insights into basic cellular processes and may lead to the discovery of novel therapeutics targeting infectious and inflammatory diseases.
Figures
{kind=link}
References
-
- Fauci AS, Morens DM. The perpetual challenge of infectious diseases. N Engl J Med. 2012;366:454–461. - PubMed
-
- Price JV, Vance RE. The macrophage paradox. Immunity. 2014;41:685–693. - PubMed
-
- Franco MP, Mulder M, Gilman RH, Smits HL. Human brucellosis. Lancet Infect Dis. 2007;7:775–786. - PubMed
-
- Piggott JA, Hochholzer L. Human melioidosis. A histopathologic study of acute and chronic melioidosis. Arch Pathol. 1970;90:101–111. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources