GLT-1-Dependent Disruption of CNS Glutamate Homeostasis and Neuronal Function by the Protozoan Parasite Toxoplasma gondii
- PMID: 27281462
- PMCID: PMC4900626
- DOI: 10.1371/journal.ppat.1005643
GLT-1-Dependent Disruption of CNS Glutamate Homeostasis and Neuronal Function by the Protozoan Parasite Toxoplasma gondii
Abstract
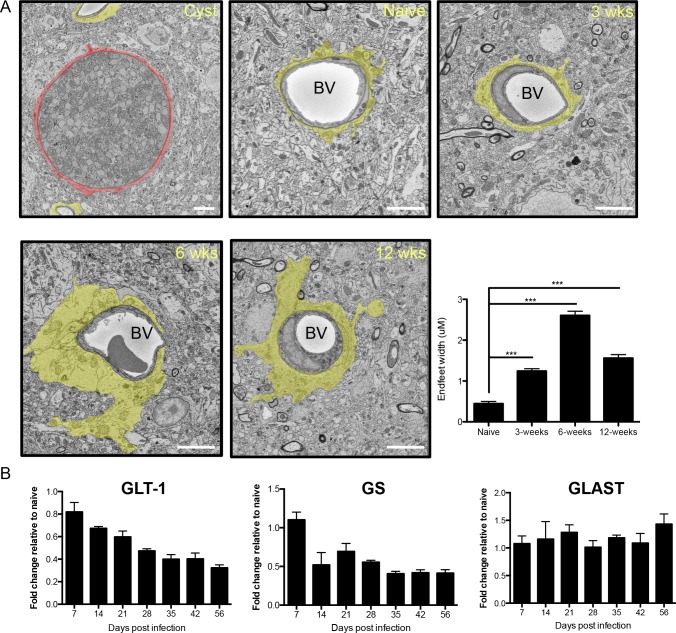
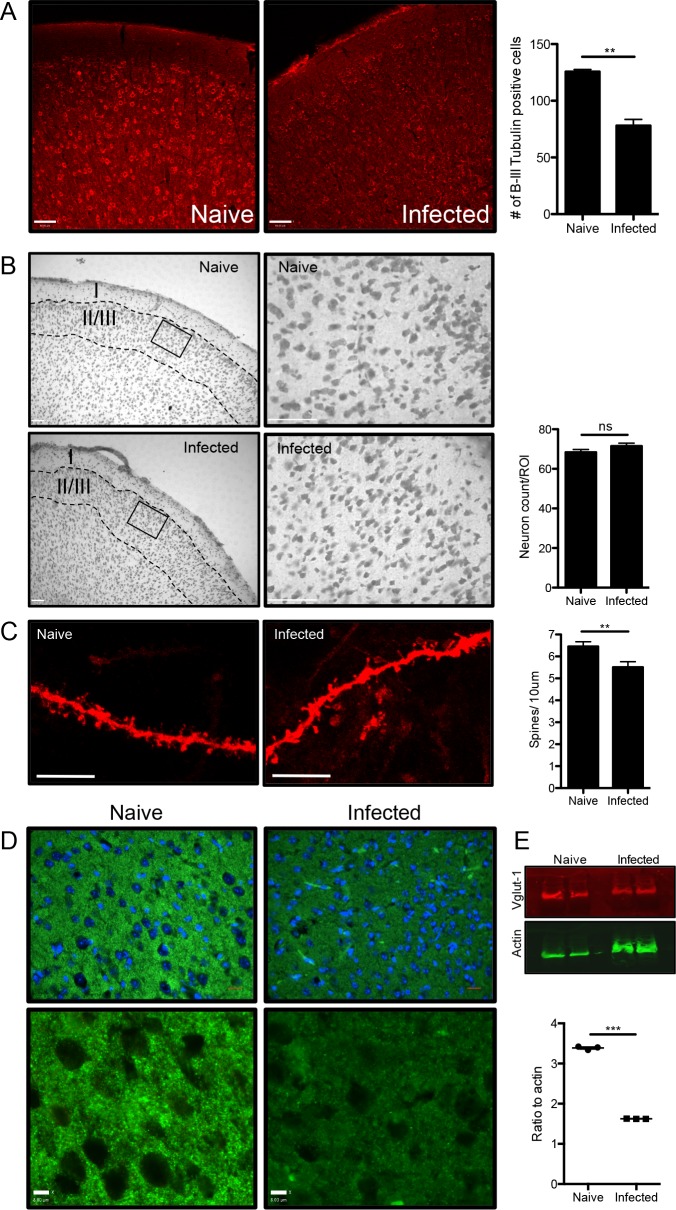
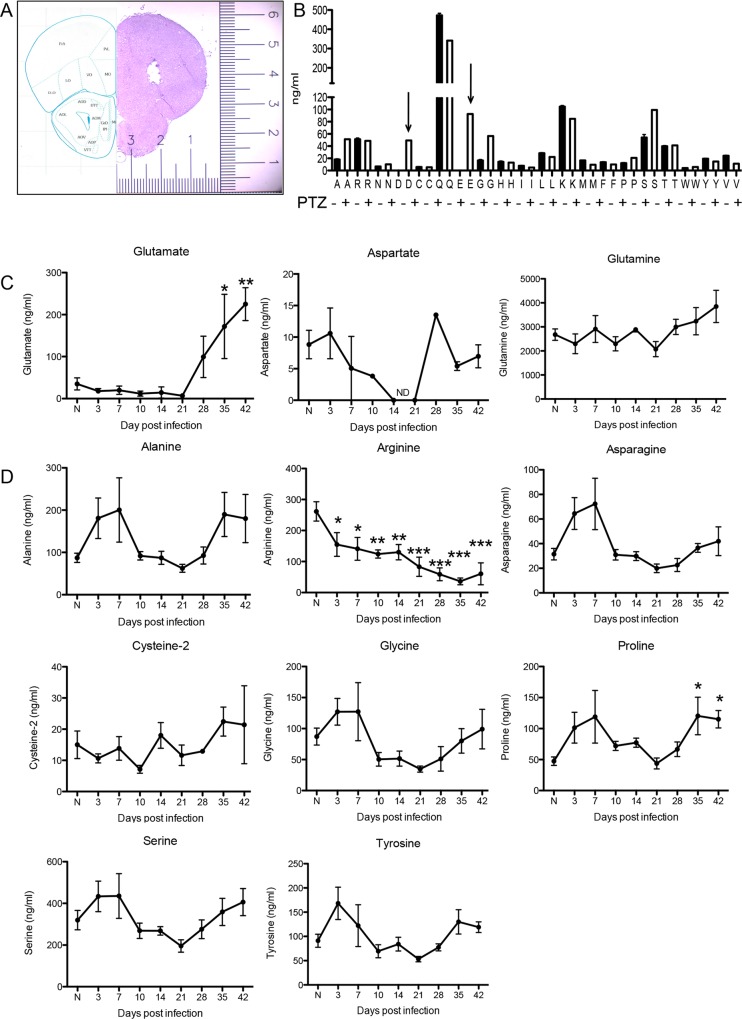
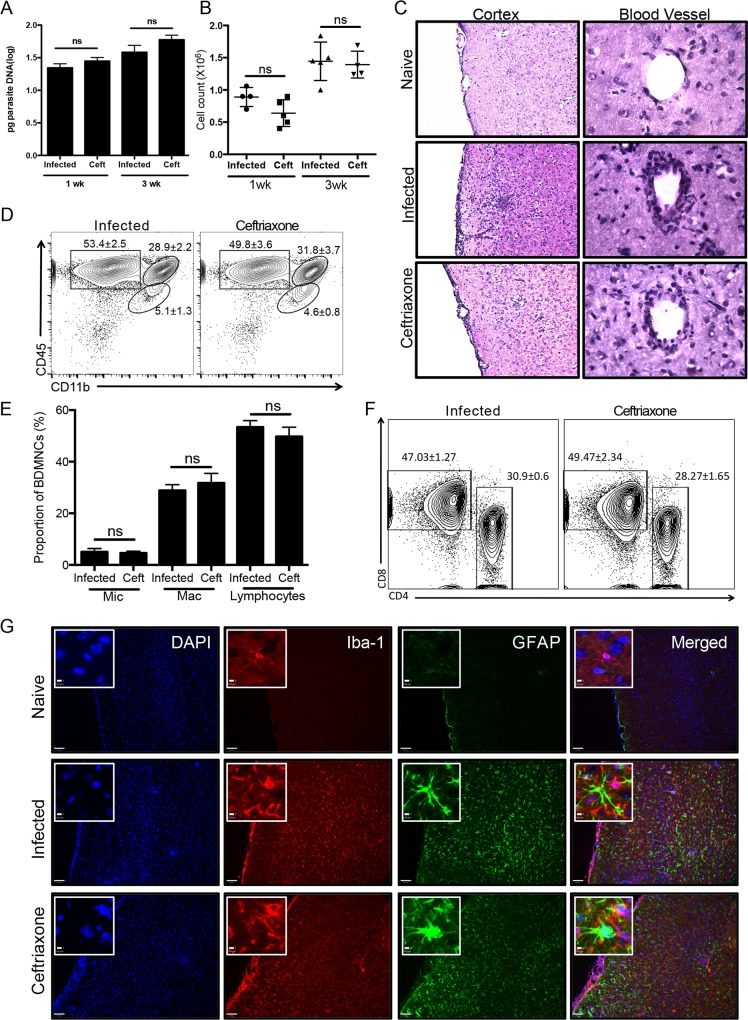
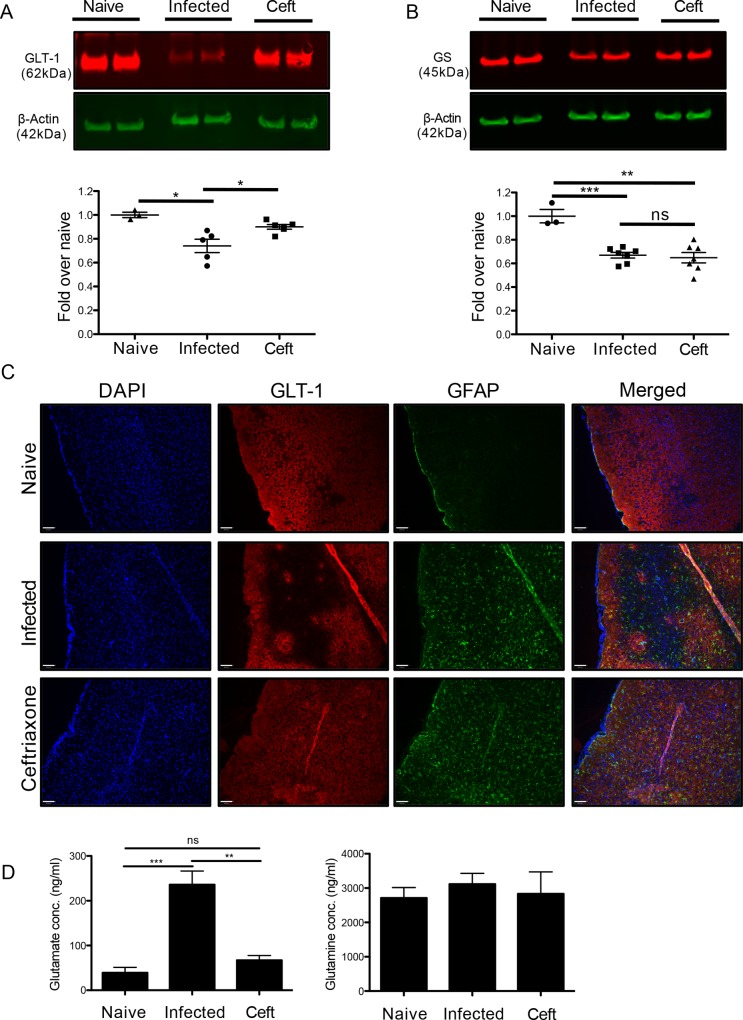
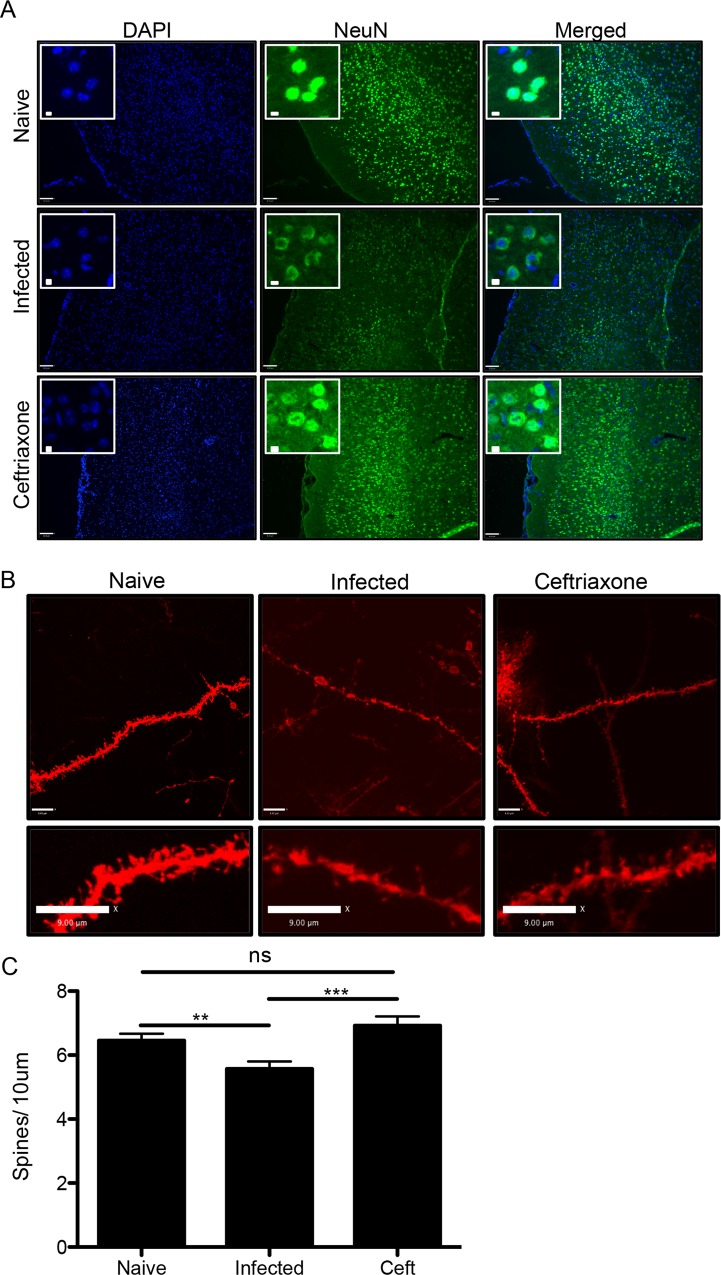
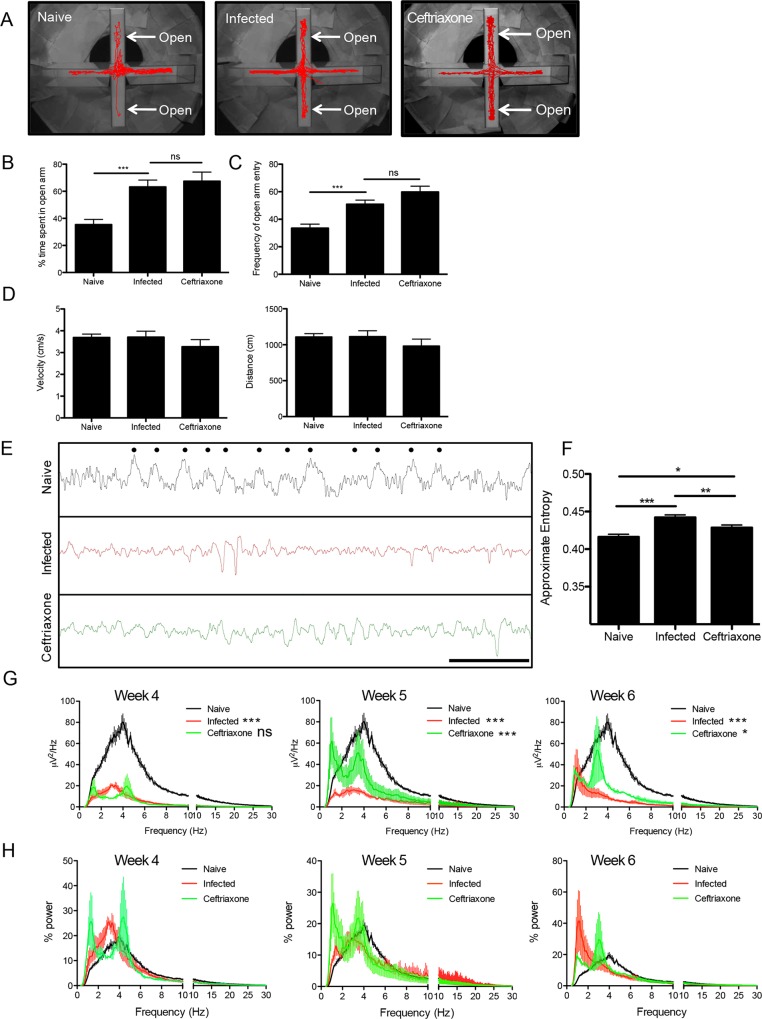
The immune privileged nature of the CNS can make it vulnerable to chronic and latent infections. Little is known about the effects of lifelong brain infections, and thus inflammation, on the neurological health of the host. Toxoplasma gondii is a parasite that can infect any mammalian nucleated cell with average worldwide seroprevalence rates of 30%. Infection by Toxoplasma is characterized by the lifelong presence of parasitic cysts within neurons in the brain, requiring a competent immune system to prevent parasite reactivation and encephalitis. In the immunocompetent individual, Toxoplasma infection is largely asymptomatic, however many recent studies suggest a strong correlation with certain neurodegenerative and psychiatric disorders. Here, we demonstrate a significant reduction in the primary astrocytic glutamate transporter, GLT-1, following infection with Toxoplasma. Using microdialysis of the murine frontal cortex over the course of infection, a significant increase in extracellular concentrations of glutamate is observed. Consistent with glutamate dysregulation, analysis of neurons reveal changes in morphology including a reduction in dendritic spines, VGlut1 and NeuN immunoreactivity. Furthermore, behavioral testing and EEG recordings point to significant changes in neuronal output. Finally, these changes in neuronal connectivity are dependent on infection-induced downregulation of GLT-1 as treatment with the ß-lactam antibiotic ceftriaxone, rescues extracellular glutamate concentrations, neuronal pathology and function. Altogether, these data demonstrate that following an infection with T. gondii, the delicate regulation of glutamate by astrocytes is disrupted and accounts for a range of deficits observed in chronic infection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Fagard R, Tan HV, Creuzet C, Pelloux H. References: Differential Development of Toxoplasma gondii in Neural Cells. Parasitology. 1999;15. - PubMed
-
- Gazzinelli R, Xu Y, Hieny S, Cheever a, Sher a. Simultaneous depletion of CD4+ and CD8+ T lymphocytes is required to reactivate chronic infection with Toxoplasma gondii. Journal of immunology (Baltimore, Md: 1950). 1992;149:175–80. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases