Broadly neutralizing epitopes in the Plasmodium vivax vaccine candidate Duffy Binding Protein
- PMID: 27194724
- PMCID: PMC4896725
- DOI: 10.1073/pnas.1600488113
Broadly neutralizing epitopes in the Plasmodium vivax vaccine candidate Duffy Binding Protein
Abstract
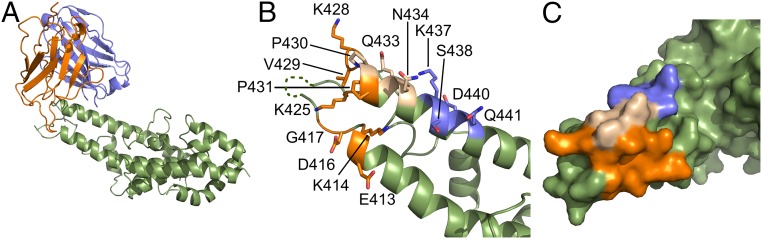
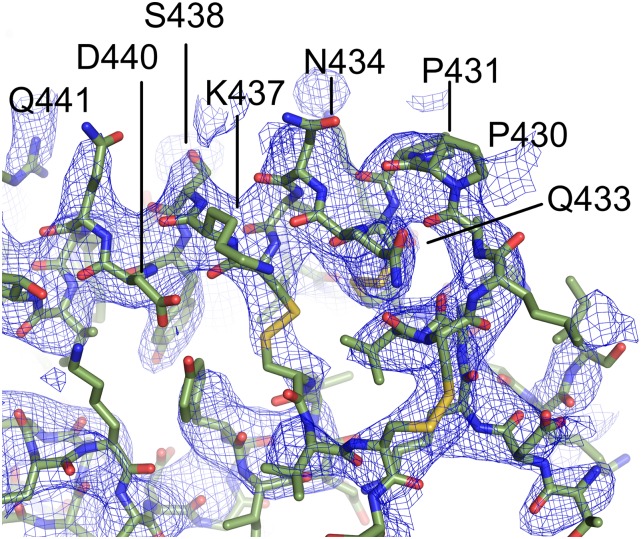
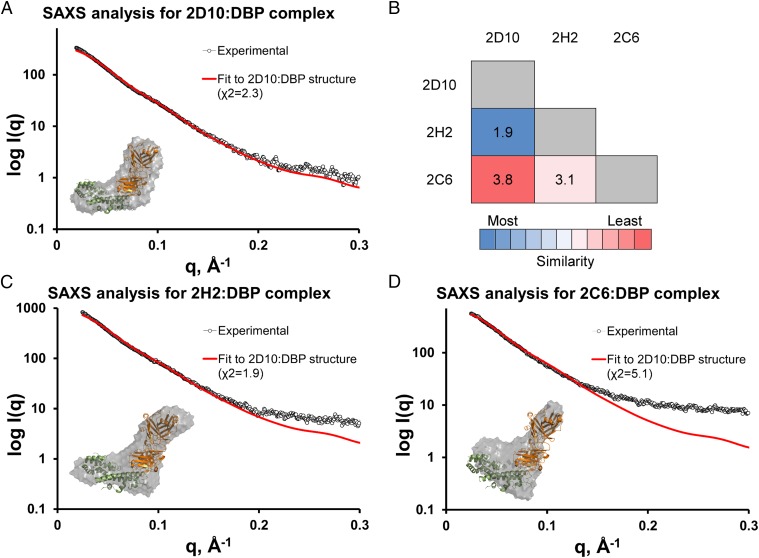
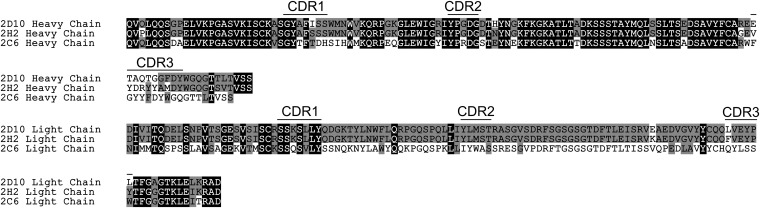
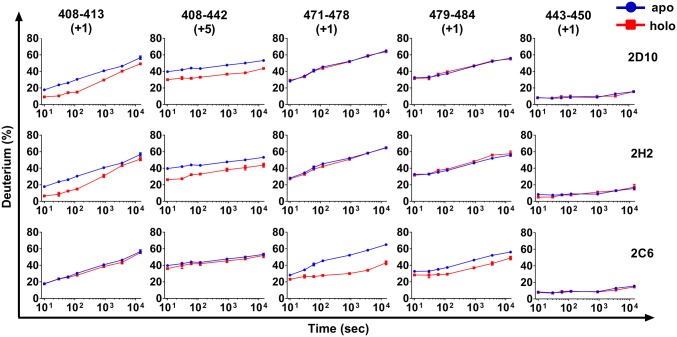
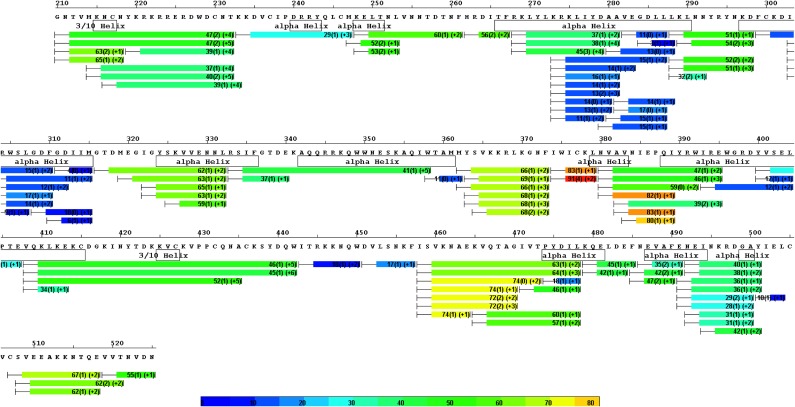
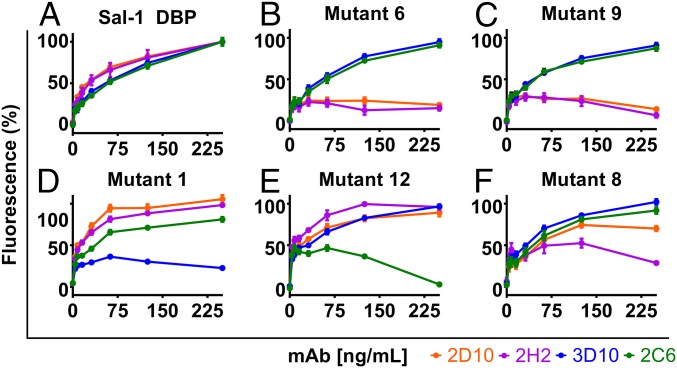
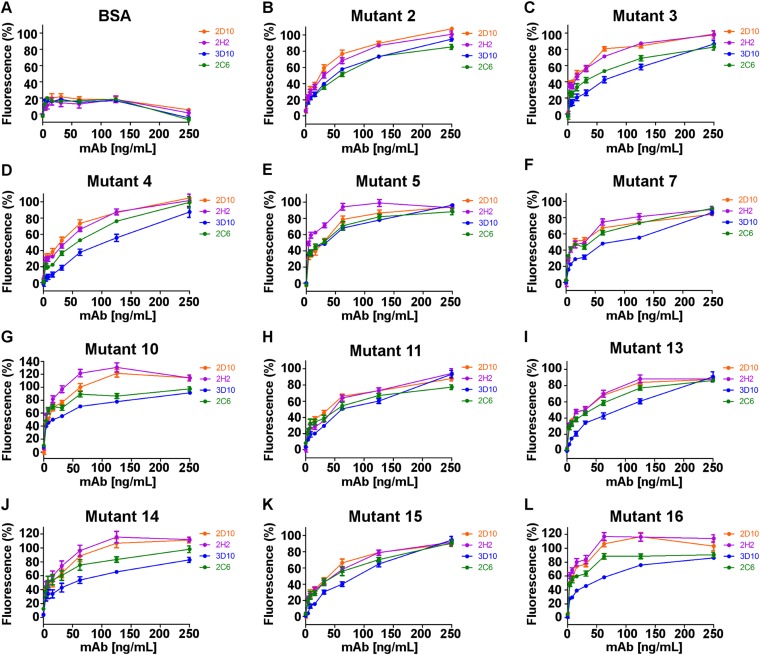
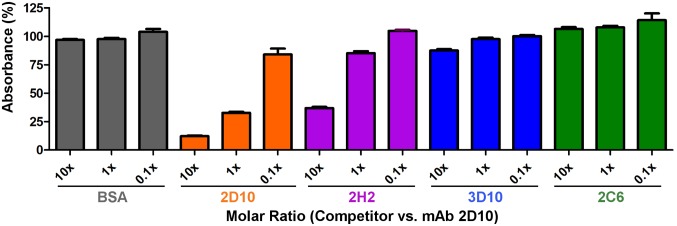
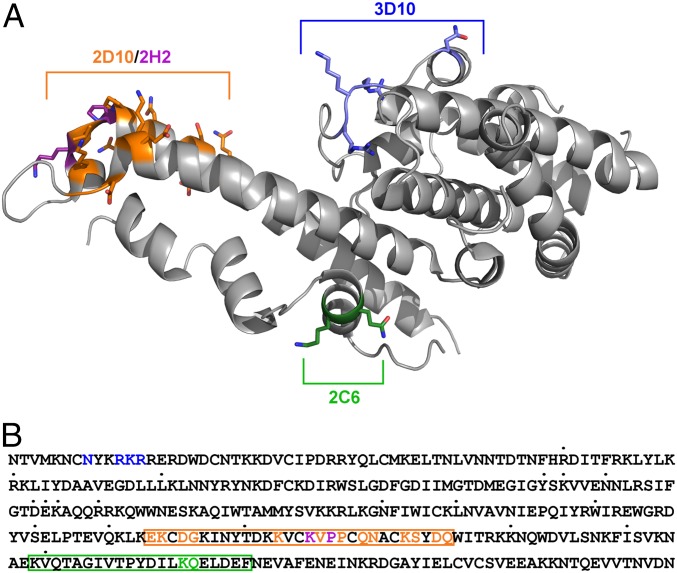
Plasmodium vivax Duffy Binding Protein (PvDBP) is the most promising vaccine candidate for P. vivax malaria. The polymorphic nature of PvDBP induces strain-specific immune responses, however, and the epitopes of broadly neutralizing antibodies are unknown. These features hamper the rational design of potent DBP-based vaccines and necessitate the identification of globally conserved epitopes. Using X-ray crystallography, small-angle X-ray scattering, hydrogen-deuterium exchange mass spectrometry, and mutational mapping, we have defined epitopes for three inhibitory mAbs (mAbs 2D10, 2H2, and 2C6) and one noninhibitory mAb (3D10) that engage DBP. These studies expand the currently known inhibitory epitope repertoire by establishing protective motifs in subdomain three outside the receptor-binding and dimerization residues of DBP, and introduce globally conserved protective targets. All of the epitopes are highly conserved among DBP alleles. The identification of broadly conserved epitopes of inhibitory antibodies provides critical motifs that should be retained in the next generation of potent vaccines for P. vivax malaria.
Keywords: Duffy Binding Protein; Plasmodium vivax; broadly neutralizing; epitopes; malaria.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources