Immune activation of the host cell induces drug tolerance in Mycobacterium tuberculosis both in vitro and in vivo
- PMID: 27114608
- PMCID: PMC4854729
- DOI: 10.1084/jem.20151248
Immune activation of the host cell induces drug tolerance in Mycobacterium tuberculosis both in vitro and in vivo
Abstract
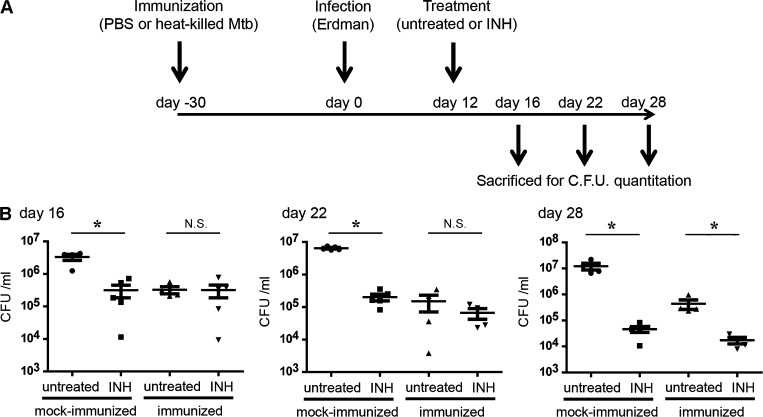
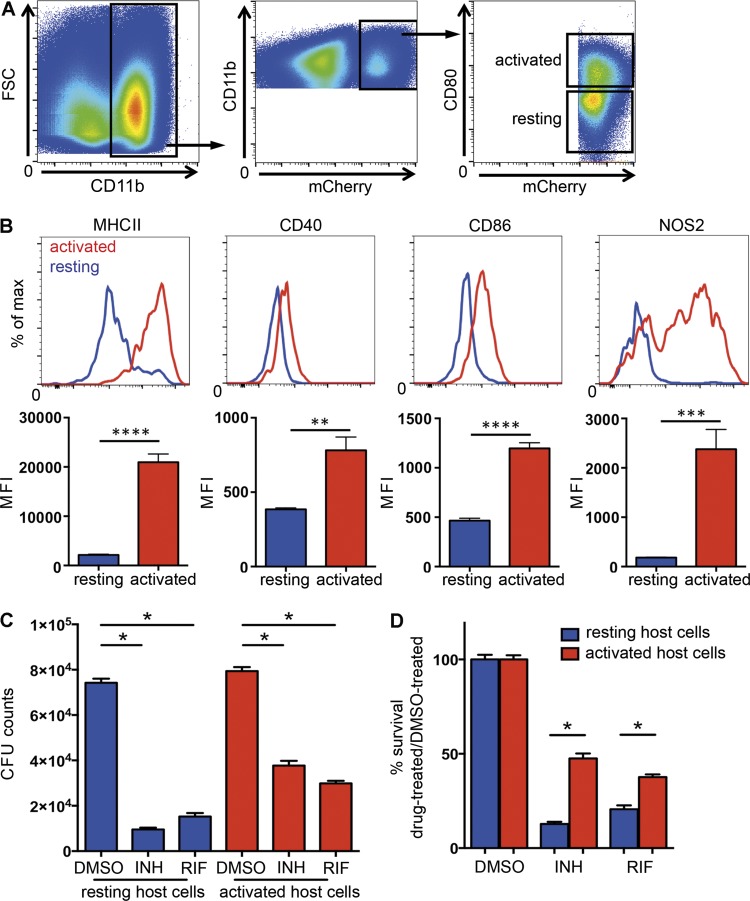
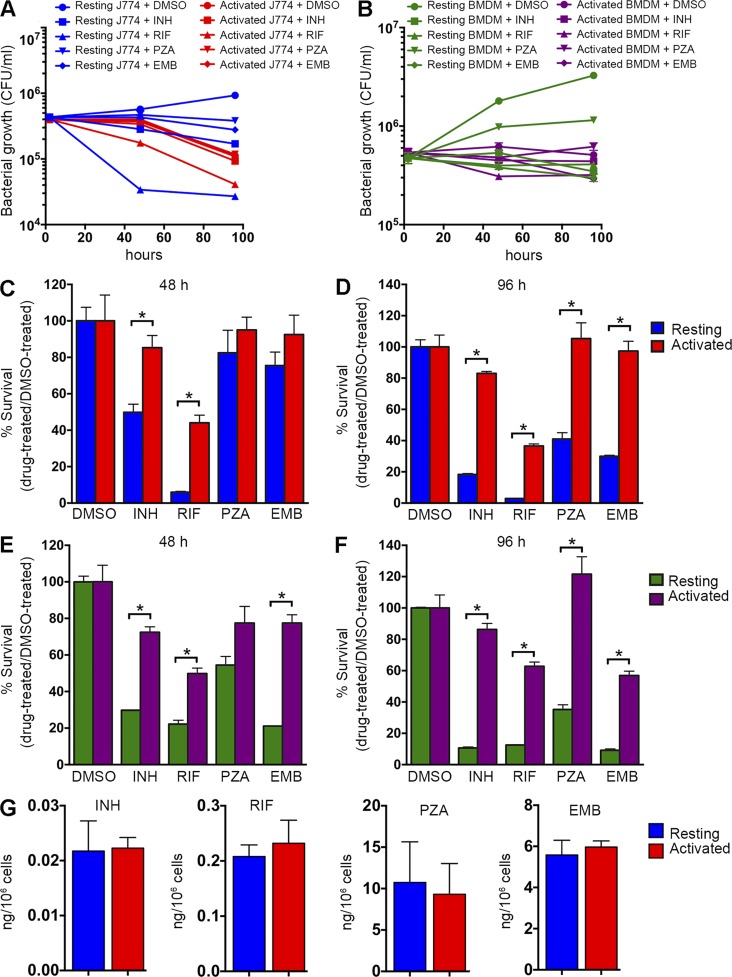
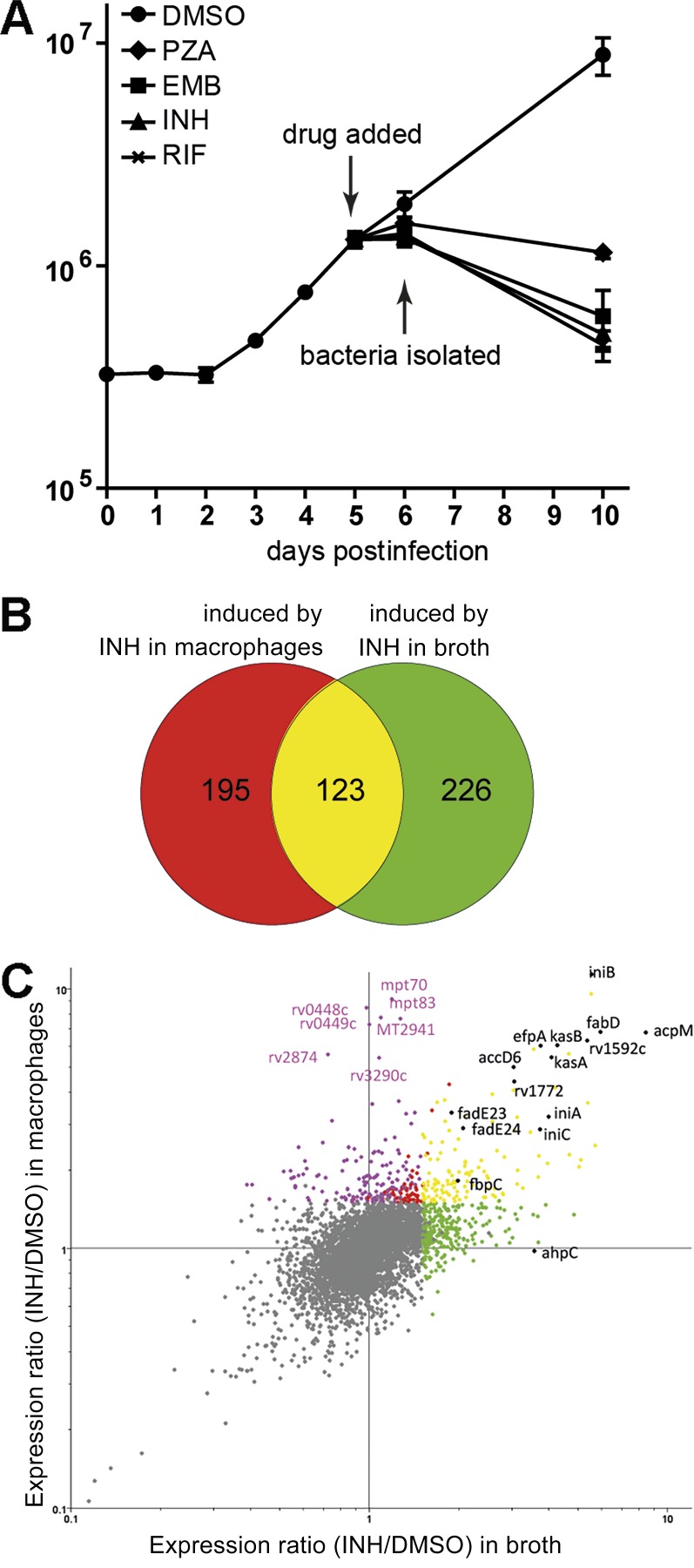
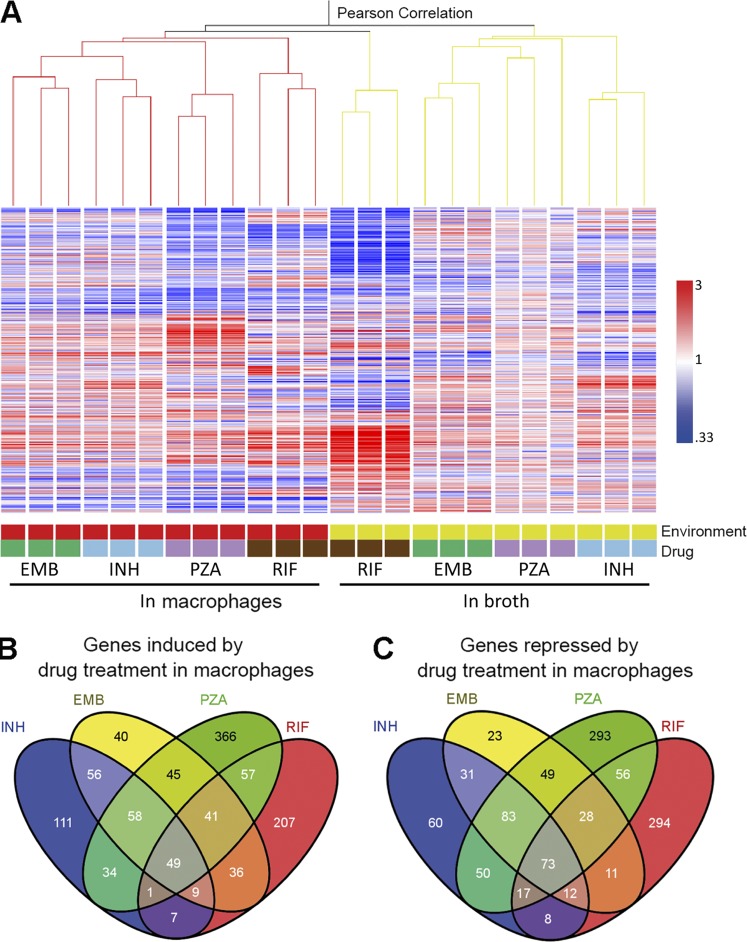
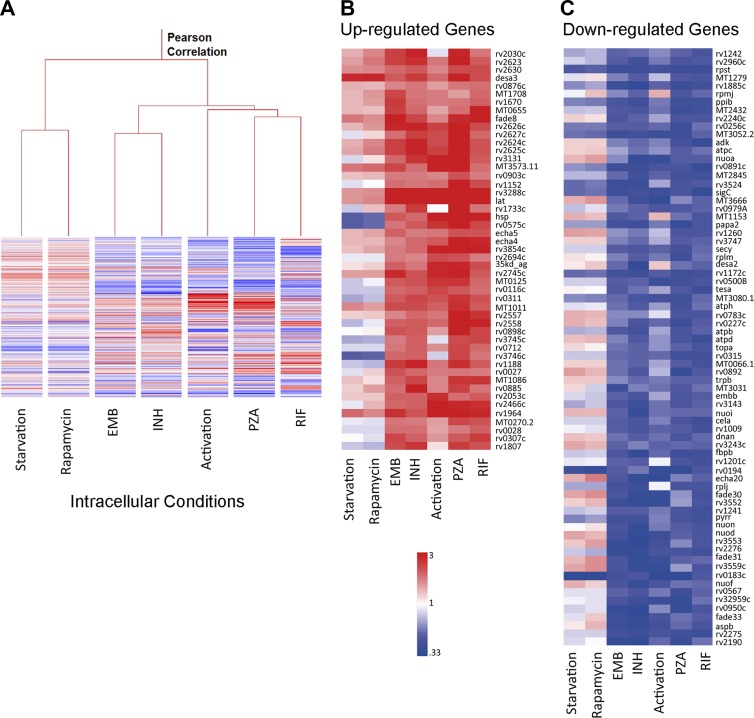
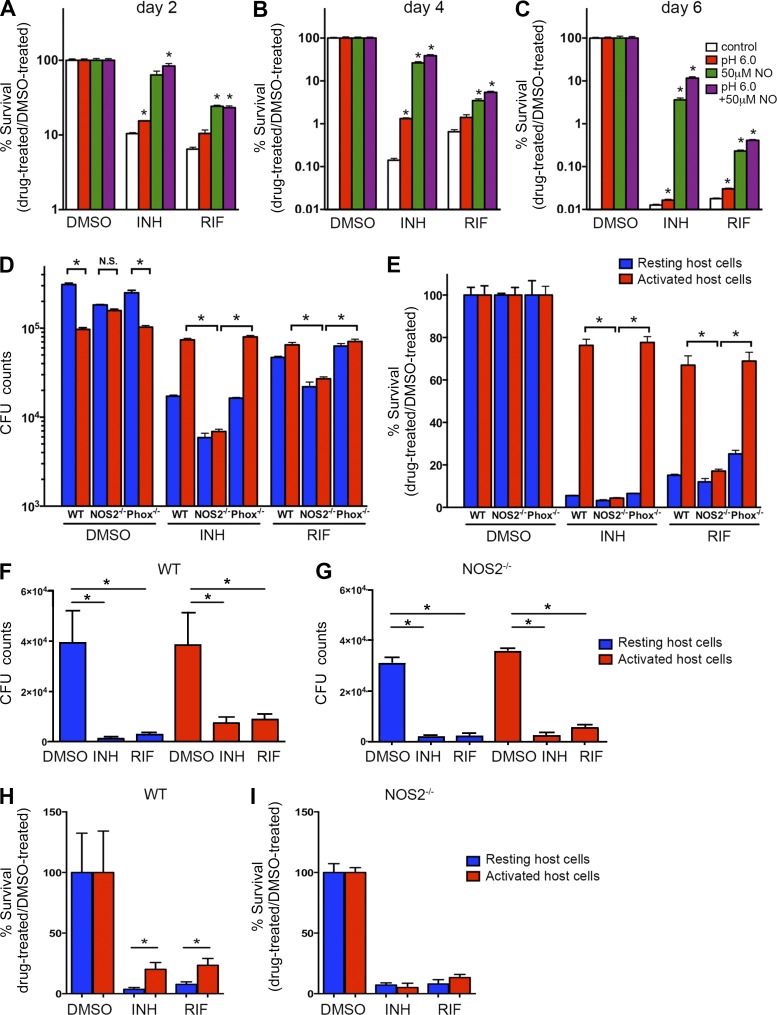
Successful chemotherapy against Mycobacterium tuberculosis (Mtb) must eradicate the bacterium within the context of its host cell. However, our understanding of the impact of this environment on antimycobacterial drug action remains incomplete. Intriguingly, we find that Mtb in myeloid cells isolated from the lungs of experimentally infected mice exhibit tolerance to both isoniazid and rifampin to a degree proportional to the activation status of the host cells. These data are confirmed by in vitro infections of resting versus activated macrophages where cytokine-mediated activation renders Mtb tolerant to four frontline drugs. Transcriptional analysis of intracellular Mtb exposed to drugs identified a set of genes common to all four drugs. The data imply a causal linkage between a loss of fitness caused by drug action and Mtb's sensitivity to host-derived stresses. Interestingly, the environmental context exerts a more dominant impact on Mtb gene expression than the pressure on the drugs' primary targets. Mtb's stress responses to drugs resemble those mobilized after cytokine activation of the host cell. Although host-derived stresses are antimicrobial in nature, they negatively affect drug efficacy. Together, our findings demonstrate that the macrophage environment dominates Mtb's response to drug pressure and suggest novel routes for future drug discovery programs.
© 2016 Liu et al.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Alland D., Kramnik I., Weisbrod T.R., Otsubo L., Cerny R., Miller L.P., Jacobs W.R. Jr., and Bloom B.R.. 1998. Identification of differentially expressed mRNA in prokaryotic organisms by customized amplification libraries (DECAL): the effect of isoniazid on gene expression in Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA. 95:13227–13232. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases