Review
doi: 10.3390/v8040106.
Molecular Insights into Crimean-Congo Hemorrhagic Fever Virus
Affiliations
- PMID: 27110812
- PMCID: PMC4848600
- DOI: 10.3390/v8040106
Item in Clipboard
Review
Molecular Insights into Crimean-Congo Hemorrhagic Fever Virus
Marko Zivcec et al.
Viruses.
.
Display options
Format
Display options
Format
Abstract
Crimean-Congo hemorrhagic fever virus (CCHFV) is a tick-borne pathogen that causes high morbidity and mortality. Efficacy of vaccines and antivirals to treat human CCHFV infections remains limited and controversial. Research into pathology and underlying molecular mechanisms of CCHFV and other nairoviruses is limited. Significant progress has been made in our understanding of CCHFV replication and pathogenesis in the past decade. Here we review the most recent molecular advances in CCHFV-related research, and provide perspectives on future research.
Keywords: Crimean-Congo hemorrhagic fever; pathogenesis; reverse genetics; tick-borne virus; viral hemorrhagic fever.
Figures
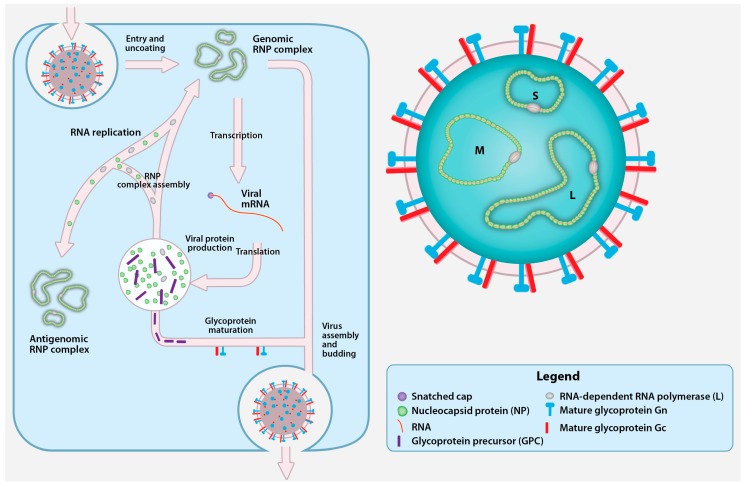{kind=link}
Crimean-Congo hemorrhagic fever virus (CCHFV) virion and replication cycle. The CCHFV virion contains a tri-segmented, negative-sense, single-stranded RNA (vRNA) genome encapsidated by the nucleoprotein (NP) and the RNA-dependent RNA polymerase (RdRp; L protein). Together, vRNA, NP, and RdRp form the genomic ribonucleoprotein complexes (RNP) inside a cellular membrane-derived envelope coated with the mature glycoproteins Gn and Gc. CCHFV attaches to an unidentified cellular receptor and enters the cells in a clathrin-dependent manner. After fusion with the cellular membrane, the viral genomic segments are uncoated and transcribed by L protein into viral mRNA that gain host cell-derived 5′ caps by cap snatching. The viral mRNAs are translated into the NP and L proteins by cytoplasmic ribosomes, while the glycoprotein precursor (GPC) appears to be translated by endoplasmic reticulum (ER)-associated ribosomes. A portion of the newly synthesized NP and L protein are used to replicate the genomic RNA by forming an RNP containing antigenomic RNA (cRNA). The GPC undergoes processing and maturation in the ER and the Golgi, and yields the Gn and Gc. Upon the accumulation of nascent mature glycoproteins and genomic RNPs, new CCHFV particles assembly is believed to occur in the Golgi followed by virion release in Golgi-derived vesicles.
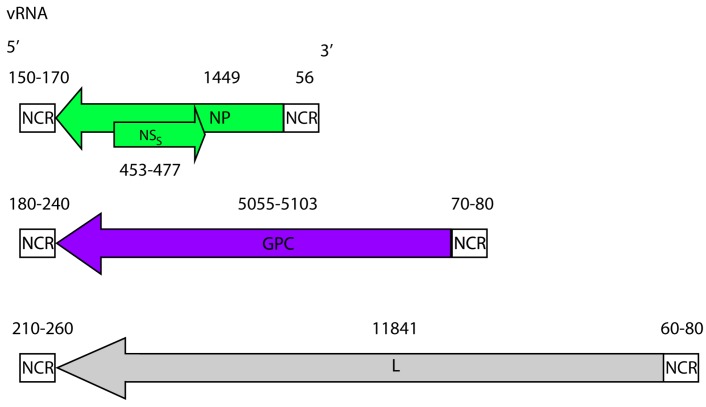{kind=link}
CCHFV genome. CCHFV possess a tri-segmented negative sense RNA genome. The small (~1.6 kb), medium (~5.4 kb) and large (~12.1 kb) segments, code for the NP, the GPC and the L protein, respectively. The small segment also codes for a non-structural S protein (NSS) in the positive sense. The coding regions are flanked by non-coding regions (NCRs). The nucleotide lengths of the regions (both coding and non-coding) are displayed and based on full-length sequences available in GenBank.
{kind=link}
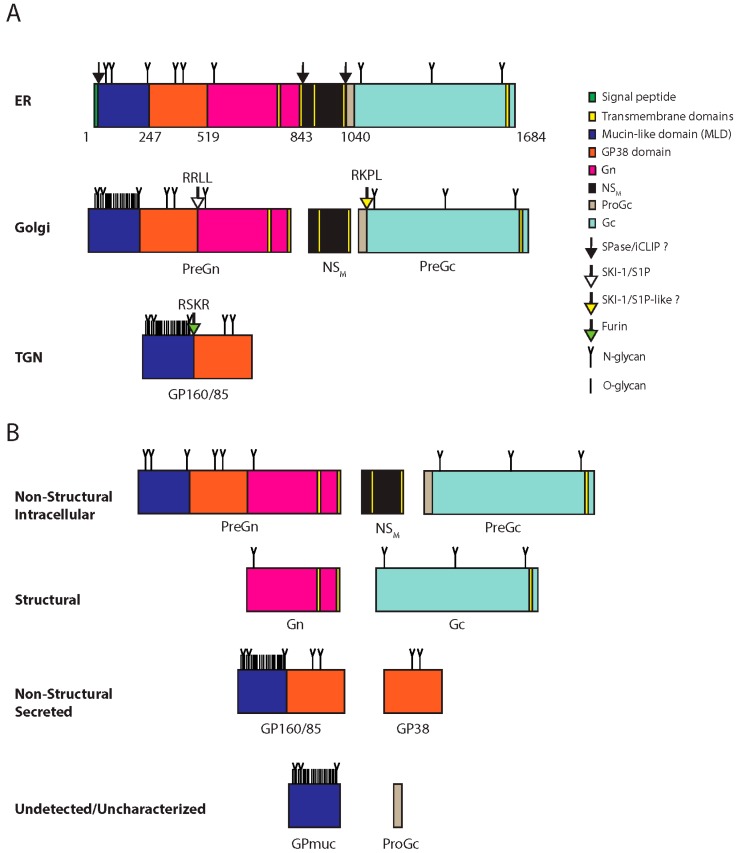
CCHFV glycoprotein processing and products. (A) CCHFV glycoprotein processing. The GPC is synthesized in the ER where N-glycosylation occurs (N-glycan). Numbers indicate amino acids positions. The signal peptidase and/or the intramembrane cleaving proteases (iCLIPs) co-translationally cleave GPC close to or within transmembrane domain-2 and -4. These cleavages yield PreGn, non-structural M protein (NSM) and PreGc. These proteins traffic to the Golgi where the mucin-like domain of PreGn is O-glycosylated (O-glycan), and is cleaved by subtilisin kexin isozyme-1/site-1 protease (SKI-1/S1P) at the RRLL motif. PreGc at the RKPL motif by a protease with similar specificity to SKI-1/SIP (SKI-1/S1P-like?). PreGn cleavage liberates an N-terminal fragment with an apparent total molecular weight on SDS-PAGE of 160 kDa (GP160) and 85 kDa (GP85). GP160/85 is later cleaved by furin in the trans-Golgi network (TGN). (B) GPC products. Processing of GPC yields non-structural products associated with the cell (Non-structural intracellular), associated with the virions (Structural), secreted but not part of the virions (Non-structural secreted), and inferred products that remain uncharacterized or have yet to be detected Undectected/uncharacterized).
{kind=link}
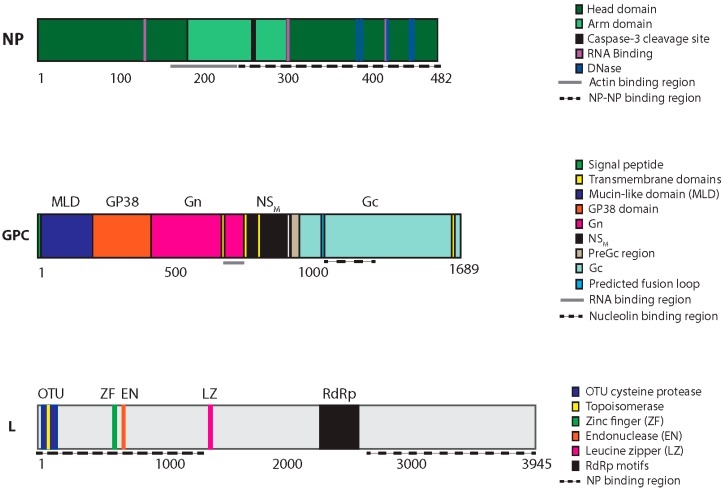
CCHFV protein domains, motifs and catalytic residues. The three CCHFV genomic segments (S, M and L) are translated into three proteins: NP, the GPC, and the L protein, respectively. The GPC is proteolytically processed to yield several additional proteins, including Gn and Gc. The approximate total size and location of motifs and catalytic residues of each protein is indicated below in amino acids.
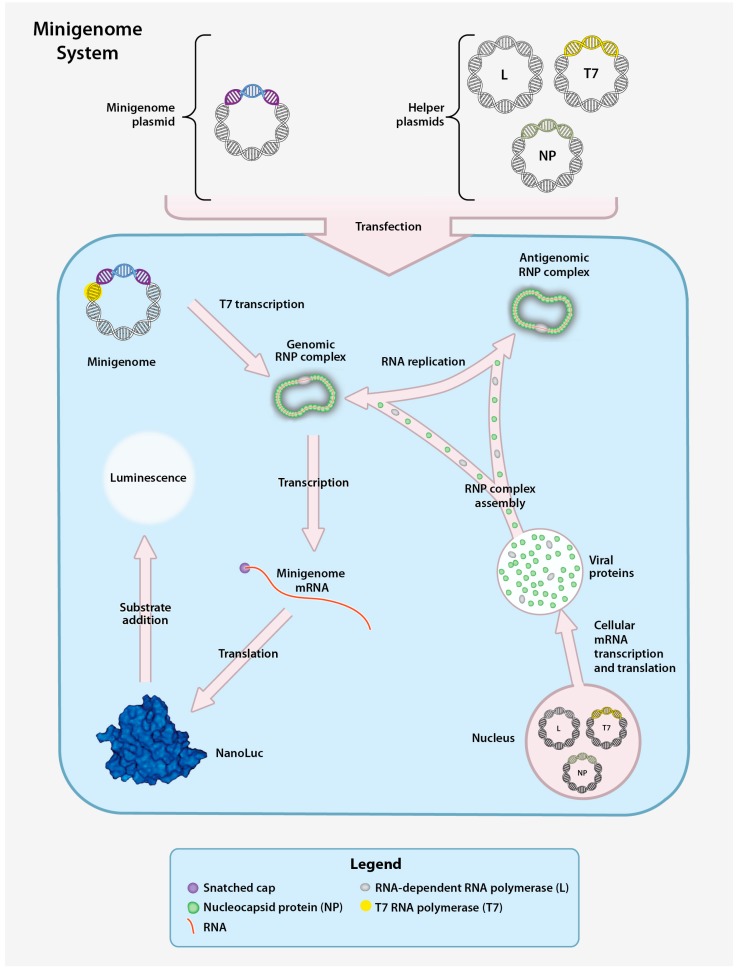{kind=link}
CCHFV minigenome system. The CCHFV minigenome system is composed of a plasmid encoding the minigenome and three helper plasmids encoding the CCHFV NP, L, and T7 RNA polymerase (T7) genes downstream of a RNA polymerase II promoter. Downstream of a T7 promoter, the minigenome plasmid contains the 5′ and 3′ non-coding regions (NCR) of a CCHFV genomic segment (S, M, or L) flanking a gene encoding a reporter protein (NanoLuc) in the negative orientation. Transfection of the helper plasmids yields the corresponding proteins to enable transcription of the minigenome plasmid and production of minigenome-derived vRNA. Following T7 transcription, the vRNA is encapsidated to form the genomic RNP. The RNP is subsequently transcribed into mRNA (secondary transcription) and translated to yield the reporter protein or replicated to produce additional vRNA. vRNA generated by both T7 transcription and RNA replication can be used as templates for transcription of reporter gene mRNA by NP and the L protein, resulting in enhanced reporter activity. A measurable luminescent signal is produced by hydrolysis of an externally provided reporter substrate.
{kind=link}
Transcriptionally and entry competent CCHF Virus-like-Particle (tec-VLP) system. The VLP system is composed of the minigenome plasmid and four helper plasmids that collectively encode the T7 polymerase, CCHFV NP, L, and GPC downstream of a cellular RNA polymerase II promoter. The corresponding proteins facilitate the generation of minigenome plasmid-derived vRNA. The minigenome plasmid is transcribed by T7 and the resulting vRNA is encapsidated to form the genomic RNP. The genomic vRNA is amplified from antigenomic RNPs, and subsequently transcribed and translated into the reporter protein. Reporter activity is monitored by adding a luciferase substrate and measuring the luminescent signal. VLPs assemble and bud from transfected cells and can subsequently enter other cells. Upon release of the genomic RNP from the VLP in the recipient cells, the minigenome can be amplified and transcribed into reporter mRNA, resulting in reporter protein expression.
{kind=link}
CCHFV infectious clone system. The CCHFV infectious clone system functions by co-transfection of genomic plasmids and helper plasmids into cells. The genomic plasmids each contain the DNA sequence of one of the CCHFV segments (S, M, or L) in the positive sense orientation downstream of a T7 promoter. The three helper plasmids encode the T7 RNA polymerase, the CCHFV NP and the L genes downstream of cellular RNA polymerase II promoters. Following T7 transcription of the genomic plasmid (enabled by the T7 helper plasmid), the cRNA is encapsidated by NP and L protein to form the antigenomic RNP, and is subsequently replicated by helper plasmid-derived NP and L protein. The vRNAs from the genomic RNPs are transcribed to mRNA and translated to yield CCHFV mRNA. The CCHFV mRNA are translated into additional NP, L, and GPC. The GPC undergoes processing and to yield the mature Gn and Gc. Following the accumulation of genomic RNPs and mature glycoproteins, infectious CCHFV particles assemble and are released.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous