Poly(A)-binding proteins are required for microRNA-mediated silencing and to promote target deadenylation in C. elegans
- PMID: 27095199
- PMCID: PMC4937315
- DOI: 10.1093/nar/gkw276
Poly(A)-binding proteins are required for microRNA-mediated silencing and to promote target deadenylation in C. elegans
Abstract
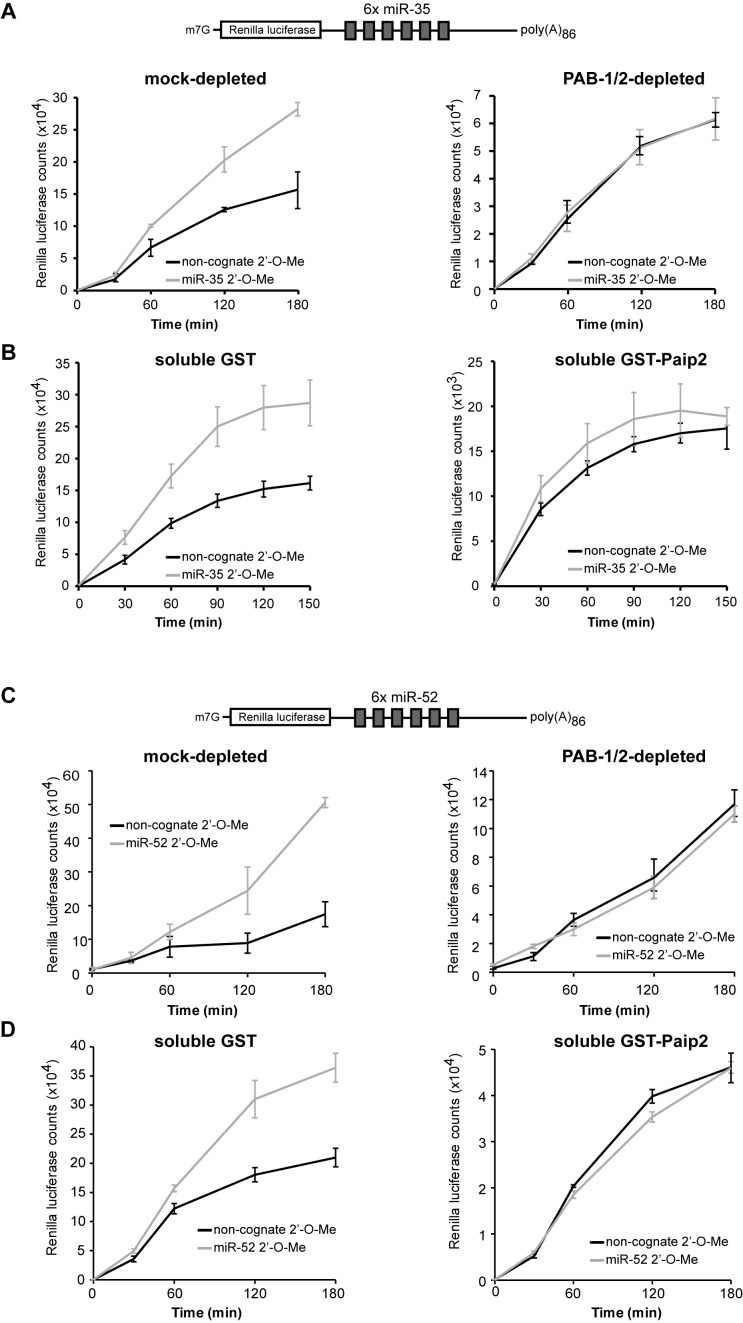
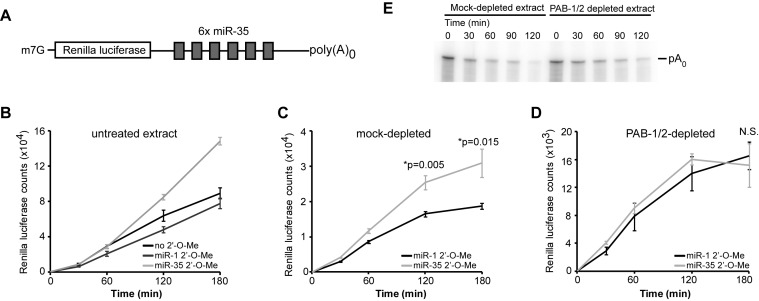
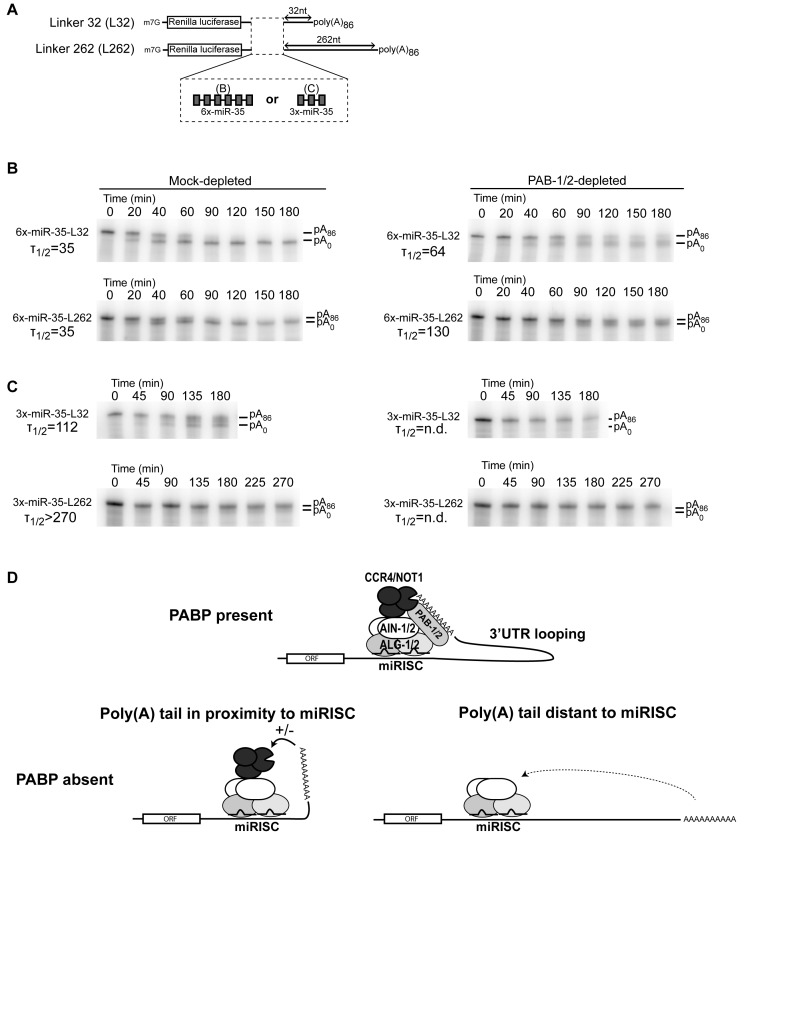
Cytoplasmic poly(A)-binding proteins (PABPs) link mRNA 3' termini to translation initiation factors, but they also play key roles in mRNA regulation and decay. Reports from mice, zebrafish and Drosophila further involved PABPs in microRNA (miRNA)-mediated silencing, but through seemingly distinct mechanisms. Here, we implicate the two Caenorhabditis elegans PABPs (PAB-1 and PAB-2) in miRNA-mediated silencing, and elucidate their mechanisms of action using concerted genetics, protein interaction analyses, and cell-free assays. We find that C. elegans PABPs are required for miRNA-mediated silencing in embryonic and larval developmental stages, where they act through a multi-faceted mechanism. Depletion of PAB-1 and PAB-2 results in loss of both poly(A)-dependent and -independent translational silencing. PABPs accelerate miRNA-mediated deadenylation, but this contribution can be modulated by 3'UTR sequences. While greater distances with the poly(A) tail exacerbate dependency on PABP for deadenylation, more potent miRNA-binding sites partially suppress this effect. Our results refine the roles of PABPs in miRNA-mediated silencing and support a model wherein they enable miRNA-binding sites by looping the 3'UTR poly(A) tail to the bound miRISC and deadenylase.
© The Author(s) 2016. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Bagga S., Bracht J., Hunter S., Massirer K., Holtz J., Eachus R., Pasquinelli A.E. Regulation by let-7 and lin-4 miRNAs results in target mRNA degradation. Cell. 2005;122:553–563. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases