Evasion of Innate Immune Responses by the Highly Virulent Cryptococcus gattii by Altering Capsule Glucuronoxylomannan Structure
- PMID: 26779451
- PMCID: PMC4701946
- DOI: 10.3389/fcimb.2015.00101
Evasion of Innate Immune Responses by the Highly Virulent Cryptococcus gattii by Altering Capsule Glucuronoxylomannan Structure
Abstract
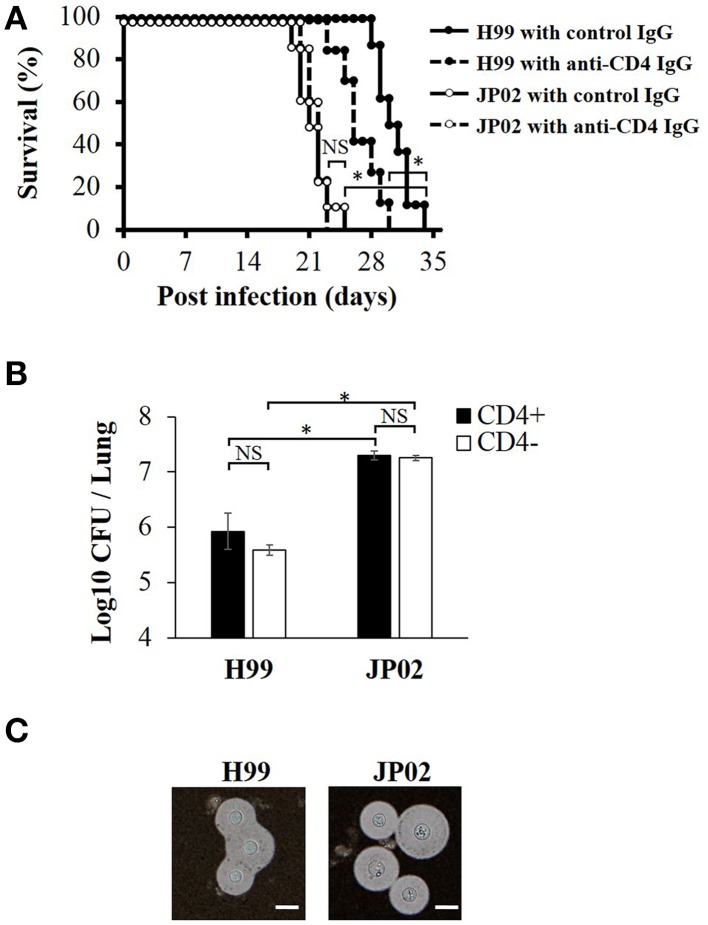
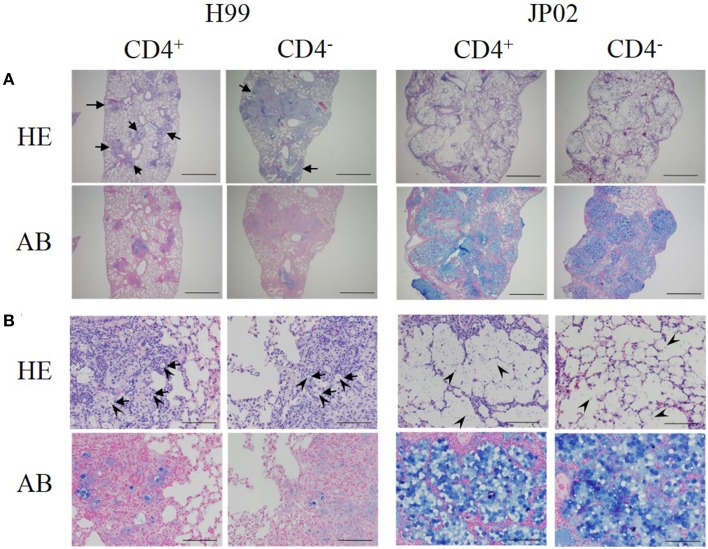
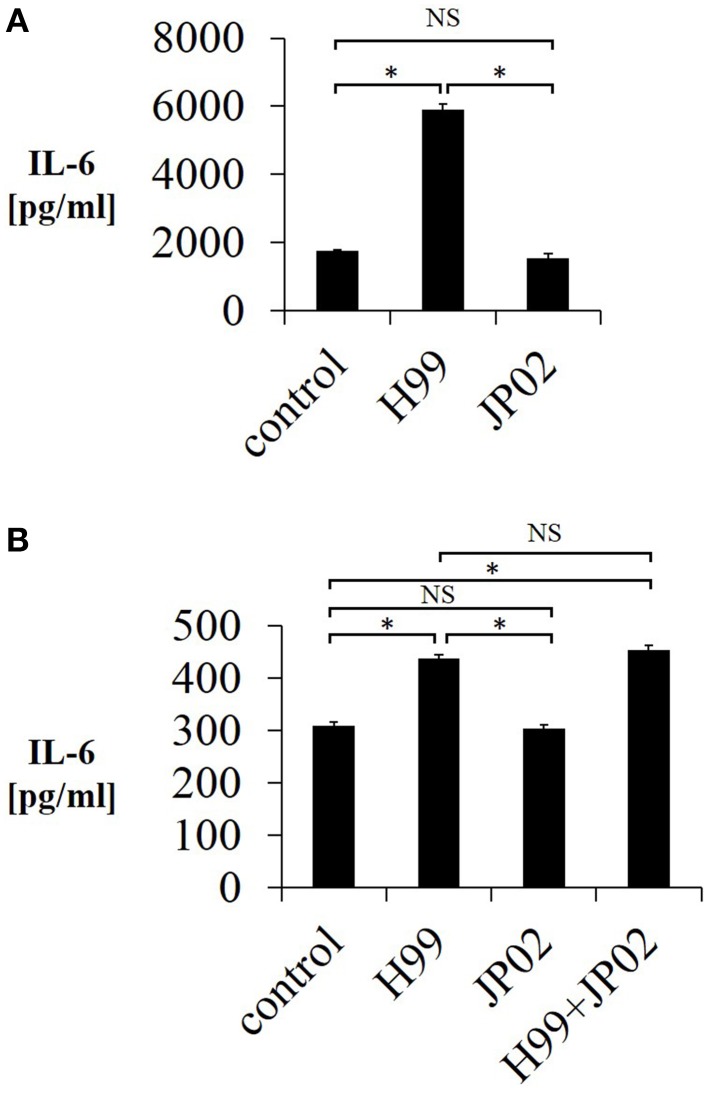
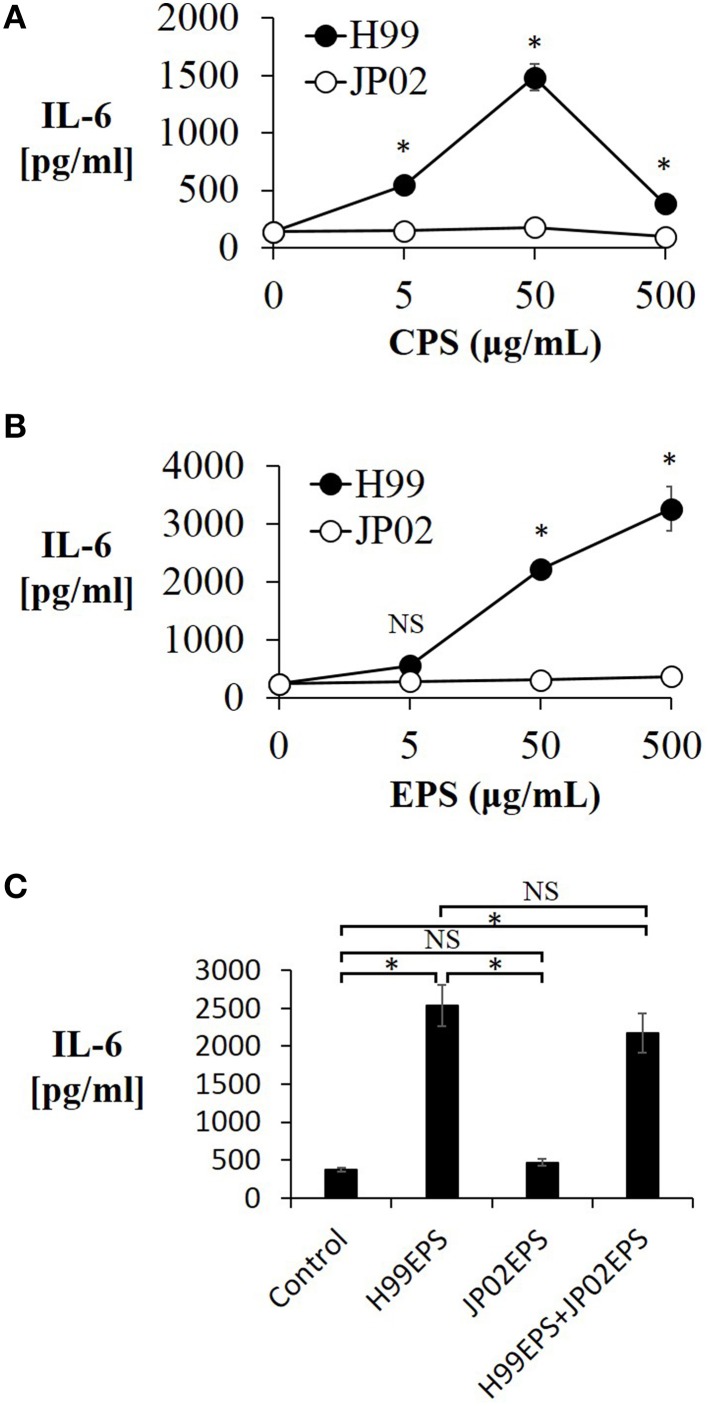
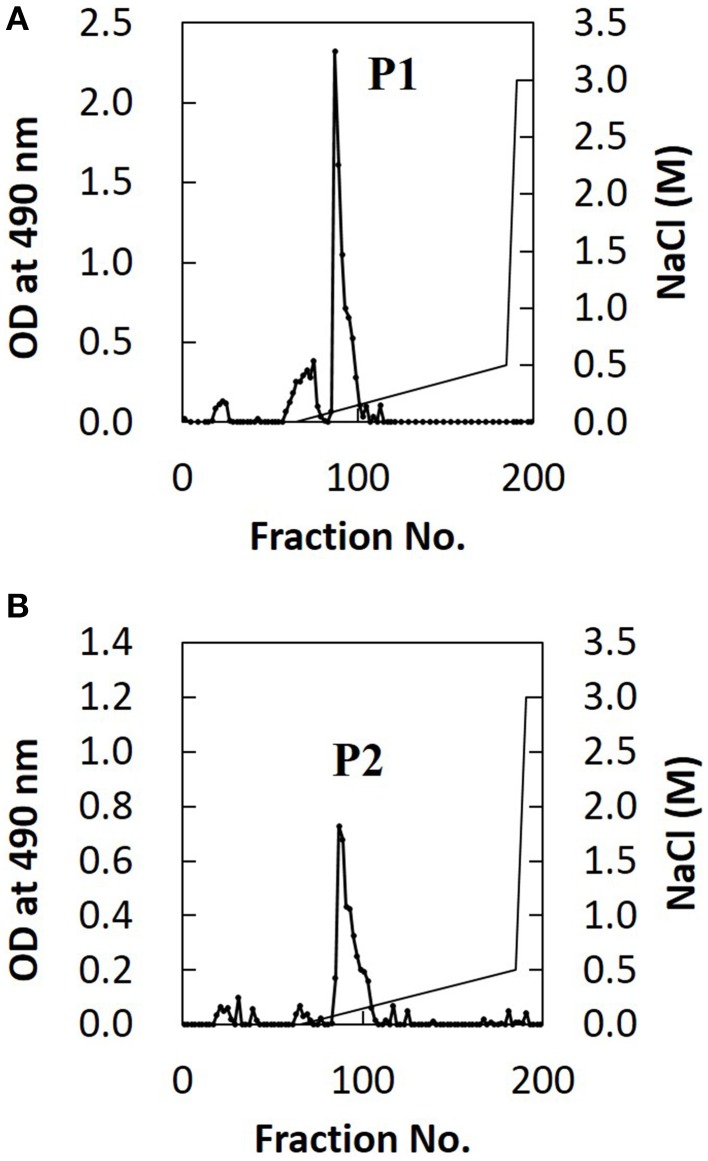
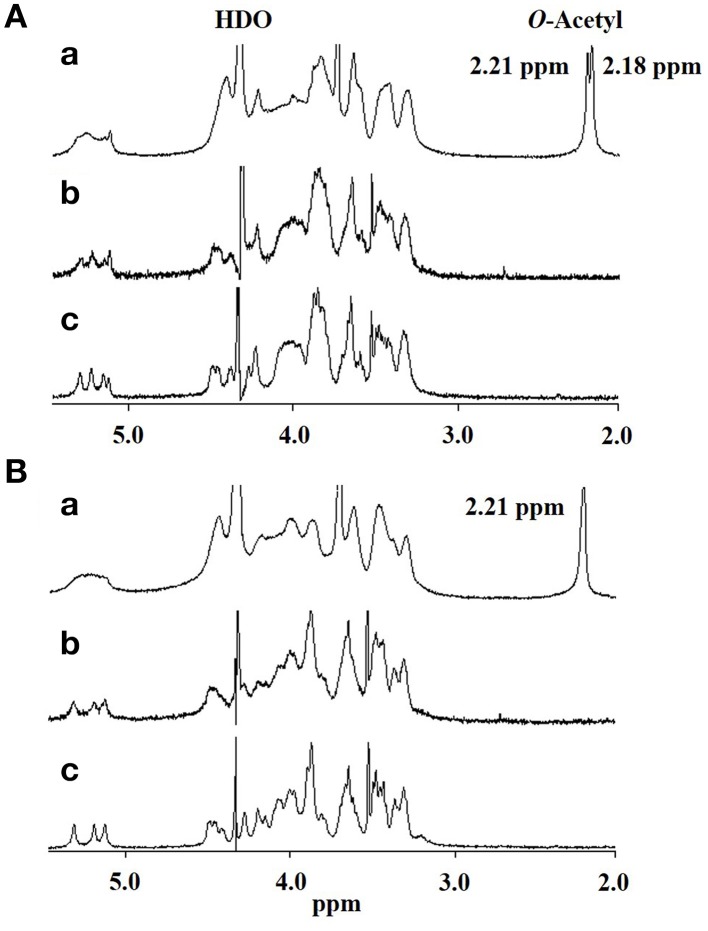
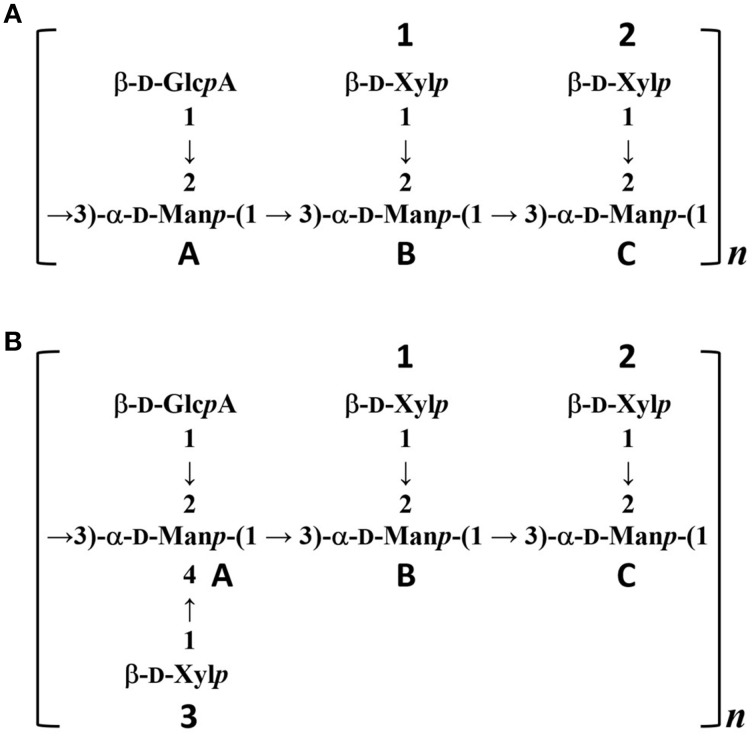
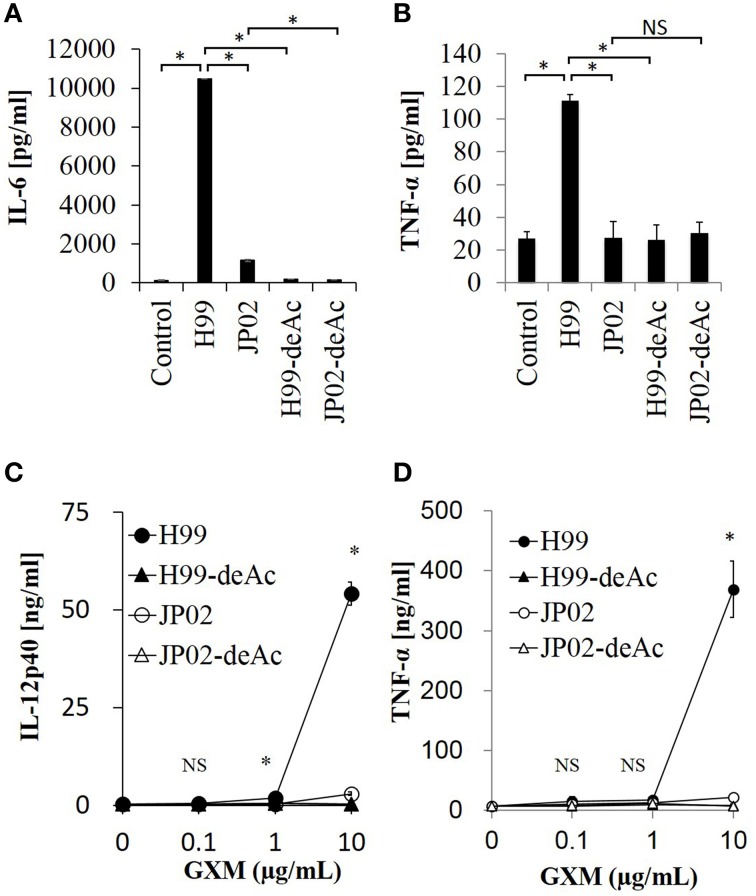
Cryptococcus neoformans causes life-threatening diseases mainly in immunosuppressed hosts such as AIDS patients; C. gattii causes disseminated infections even in healthy hosts. To identify the possible molecular mechanisms underlying this difference in virulence, we investigated the survival and histopathology of lung tissue in wild-type and CD4-depleted mice infected with C. neoformans H99 and C. gattii JP02 (the highly virulent strain isolated in Japan); we then compared dendritic cell (DC) cytokine release responses to different cell fractions from these two strains. JP02-infected mice exhibited shorter survival and fewer inflammatory cells in the lung than H99-infected control mice. Depletion of CD4-related cellular immunity reduced survival of H99-infected mice but had no effect on the survival or inflammatory cell infiltration in JP02-infected mice, suggesting that JP02 evades immune detection. To identify the molecule(s) conferring this difference, we measured cytokine production from murine DCs co-cultured with H99 and JP02 in vitro. The levels of inflammatory cytokines from DCs treated with intact JP02 cells, the extracted capsule, secreted extracellular polysaccharides, and purified glucuronoxylomannan (GXM) were markedly lower than those induced by intact H99 cells and corresponding H99 fractions. Structural analysis of GXM indicated that JP02 altered one of two O-acetyl groups detected in the H99 GXM. Deacetylated GXM lost the ability to induce inflammatory cytokine release from DCs, implicating these O-acetyl groups in immune recognition. We conclude that the highly virulent C. gattii processes a structural alteration in GXM that allows this pathogen to evade the immune response and therefore elimination.
Keywords: Cryptococcus gattii; Cryptococcus neoformans; O-acetylation; capsule; dendritic cells; glucuronoxylomannan; innate immune responses; structure.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- BC Centre for Disease Control, An agency of the Provincial Health Services Authority, British Columbia (2015). British Columbia Annual Summary of Reportable Diseases 2014. Available online at: http://www.bccdc.ca/bccdc/resources/ar2014finalsmall.pdf
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials