Environmental marine pathogen isolation using mesocosm culture of sharpsnout seabream: striking genomic and morphological features of novel Endozoicomonas sp
- PMID: 26639610
- PMCID: PMC4671022
- DOI: 10.1038/srep17609
Environmental marine pathogen isolation using mesocosm culture of sharpsnout seabream: striking genomic and morphological features of novel Endozoicomonas sp
Abstract
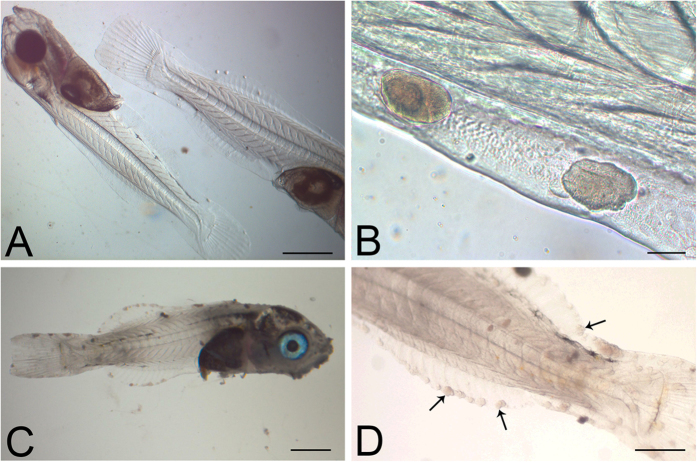
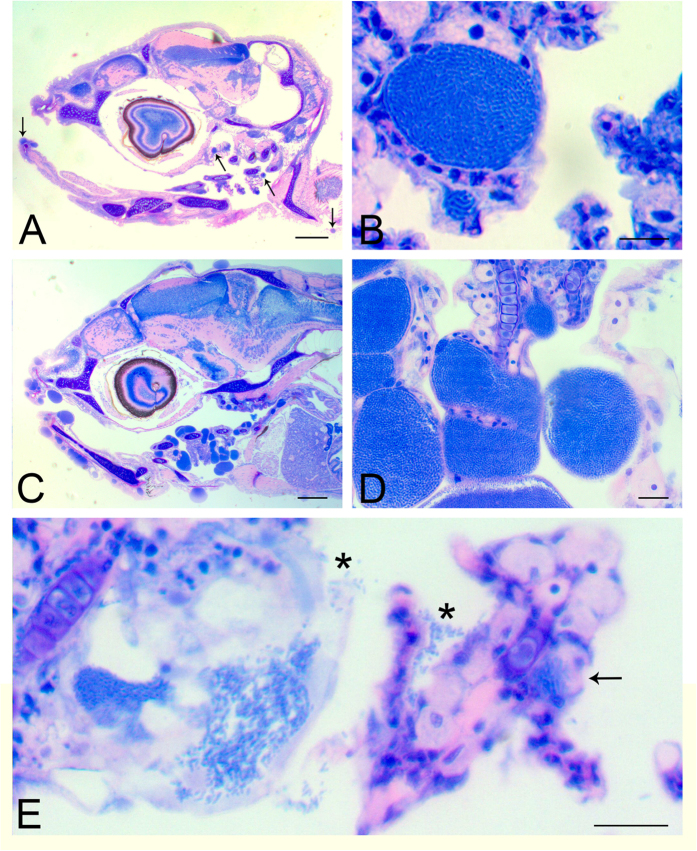
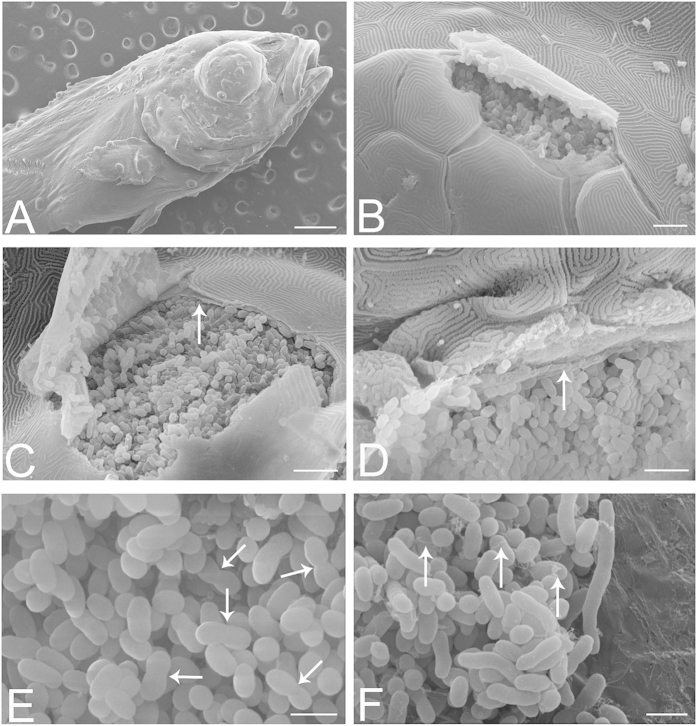
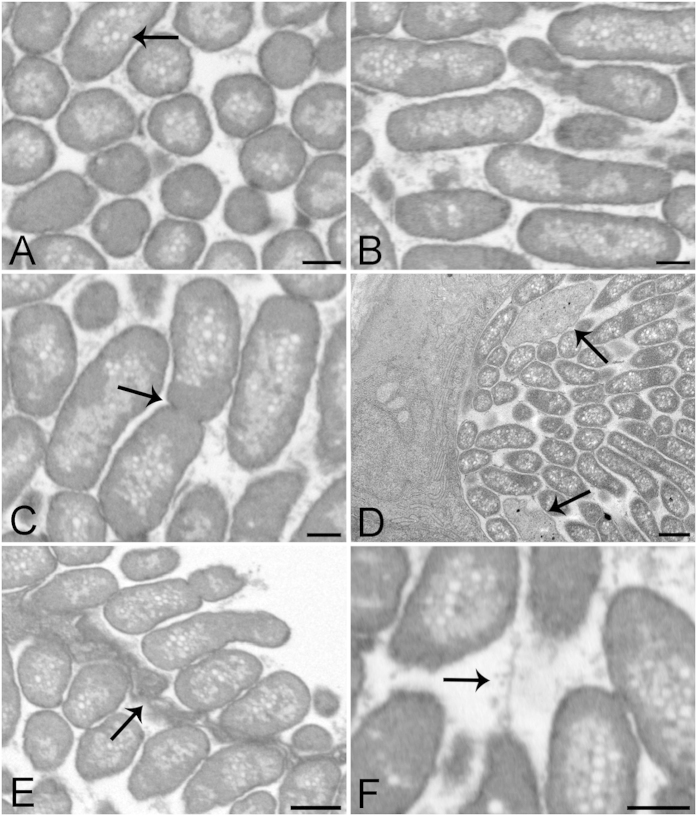
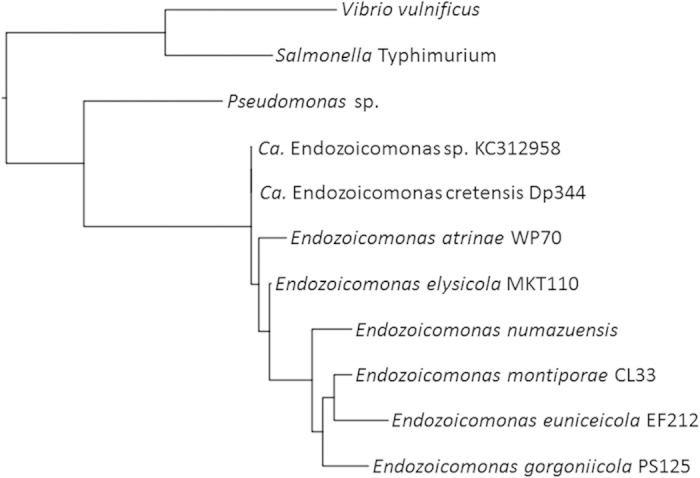
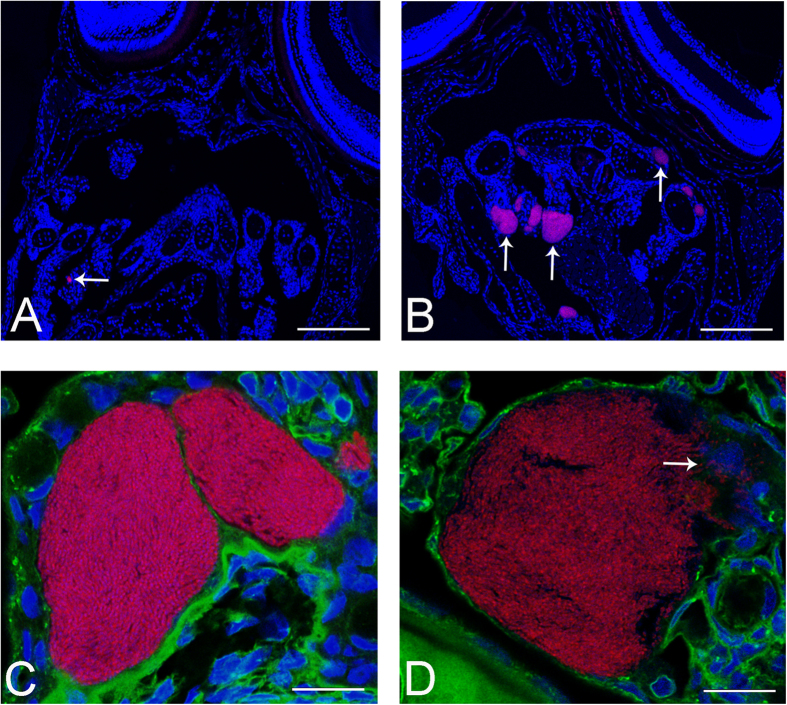
Aquaculture is a burgeoning industry, requiring diversification into new farmed species, which are often at risk from infectious disease. We used a mesocosm technique to investigate the susceptibility of sharpsnout seabream (Diplodus puntazzo) larvae to potential environmental pathogens in seawater compared to control borehole water. Fish exposed to seawater succumbed to epitheliocystis from 21 days post hatching, causing mortality in a quarter of the hosts. The pathogen responsible was not chlamydial, as is often found in epitheliocystis, but a novel species of the γ-proteobacterial genus Endozoicomonas. Detailed characterisation of this pathogen within the infectious lesions using high resolution fluorescent and electron microscopy showed densely packed rod shaped bacteria. A draft genome sequence of this uncultured bacterium was obtained from preserved material. Comparison with the genome of the Endozoicomonas elysicola type strain shows that the genome of Ca. Endozoicomonas cretensis is undergoing decay through loss of functional genes and insertion sequence expansion, often indicative of adaptation to a new niche or restriction to an alternative lifestyle. These results demonstrate the advantage of mesocosm studies for investigating the effect of environmental bacteria on susceptible hosts and provide an important insight into the genome dynamics of a novel fish pathogen.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Divinach P. & Kentouri M. Hatchery techniques for specific diversification in Mediterranean finfish larviculture. Cahiers Options Mediterraneennes 47, 75–87 (2000).
-
- Papandroulakis N., Mylonas C. C., Maingot E. & Divanach P. First results of greater amberjack (Seriola dumerili) larval rearing in mesocosm. Aquaculture 250, 155–161, 10.1016/j.aquaculture.2005年02月03日6 (2005). - DOI
-
- Papandroulakis N., Kentouri M., Maingot E. & Divanach P. Mesocosm: A Reliable Technology for Larval Rearing of Diplodus puntazzo and Diplodus sargus sargus. Aquaculture International 12, 345–355, 10.1023/B:AQUI.0000042134.21211.ab (2004). - DOI
-
- Bergh Ø. The dual myths of the healthy wild fish and the unhealthy farmed fish. Dis Aquat Org 75, 159–164 (2007). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases