Comparative Analysis of Transcriptional Profiles of Adult Schistosoma japonicum from Different Laboratory Animals and the Natural Host, Water Buffalo
- PMID: 26285138
- PMCID: PMC4540470
- DOI: 10.1371/journal.pntd.0003993
Comparative Analysis of Transcriptional Profiles of Adult Schistosoma japonicum from Different Laboratory Animals and the Natural Host, Water Buffalo
Abstract
Background: Schistosomiasis is one of the most widely distributed parasitic diseases in the world. Schistosoma japonicum, a zoonotic parasite with a wide range of mammalian hosts, is one of the major pathogens of this disease. Although numerous studies on schistosomiasis japonica have been performed using laboratory animal models, systematic comparative analysis of whole-genome expression profiles in parasites from different laboratory animals and nature mammalian hosts is lacking to date.
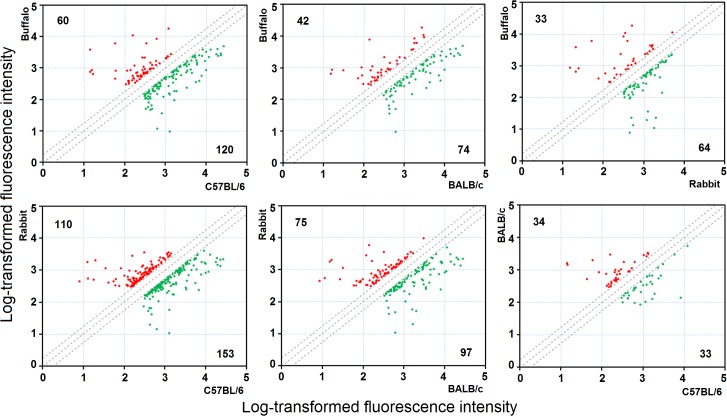
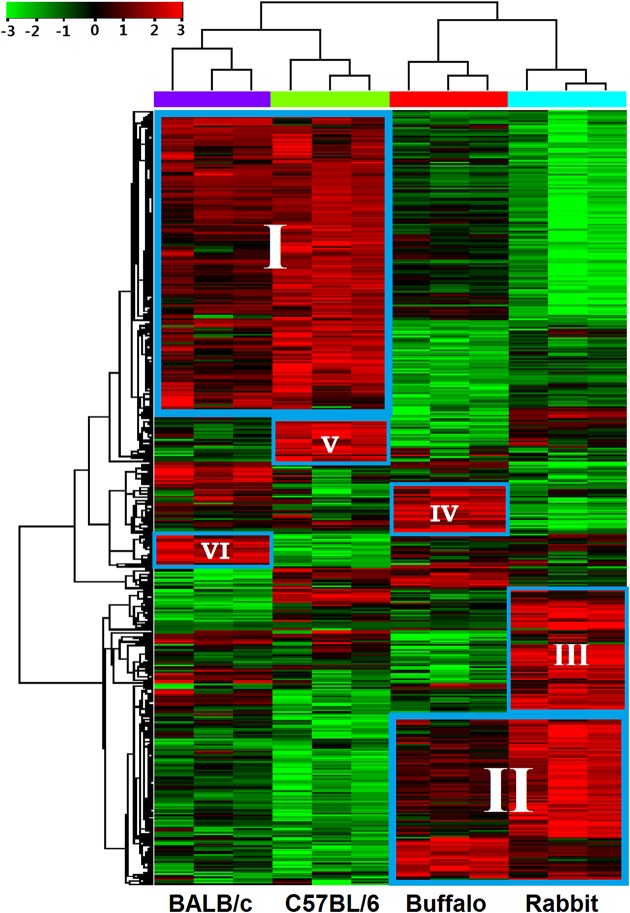
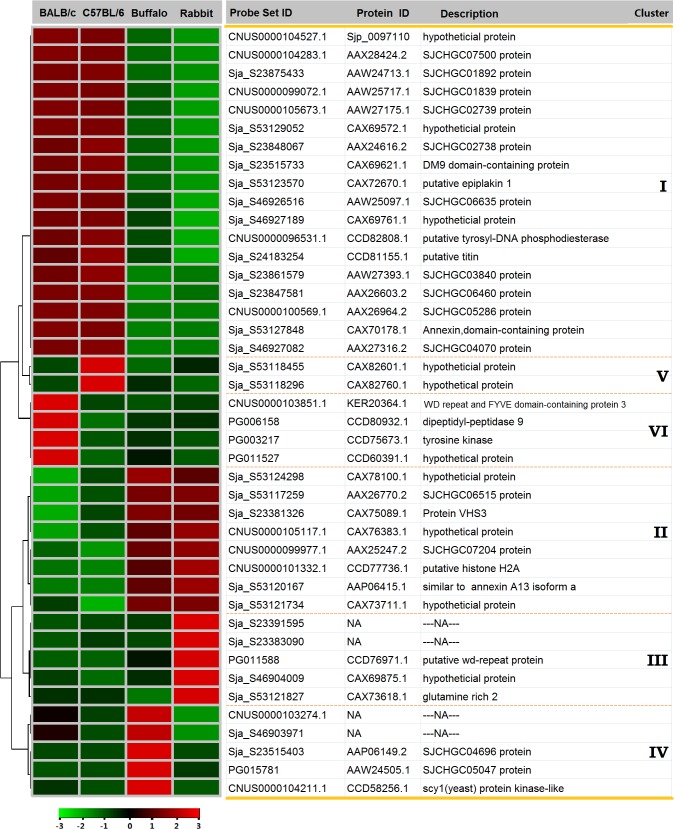
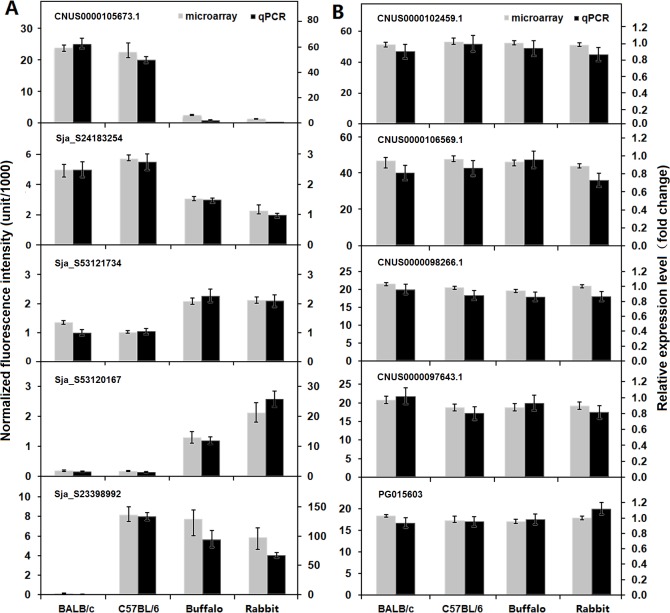
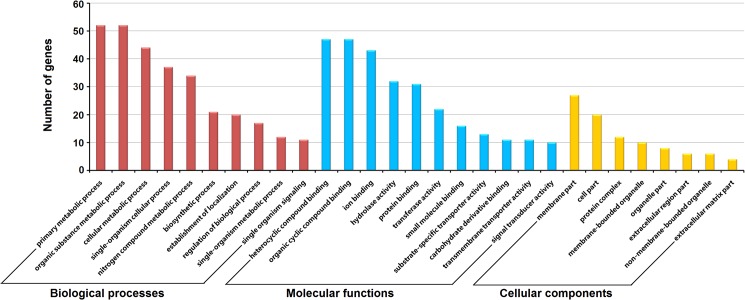
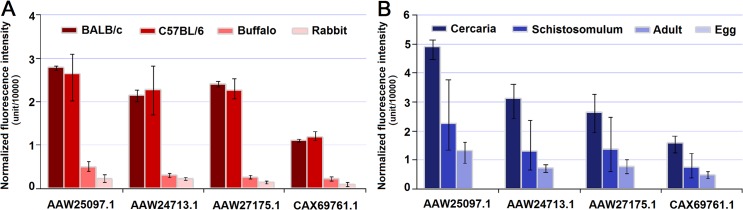
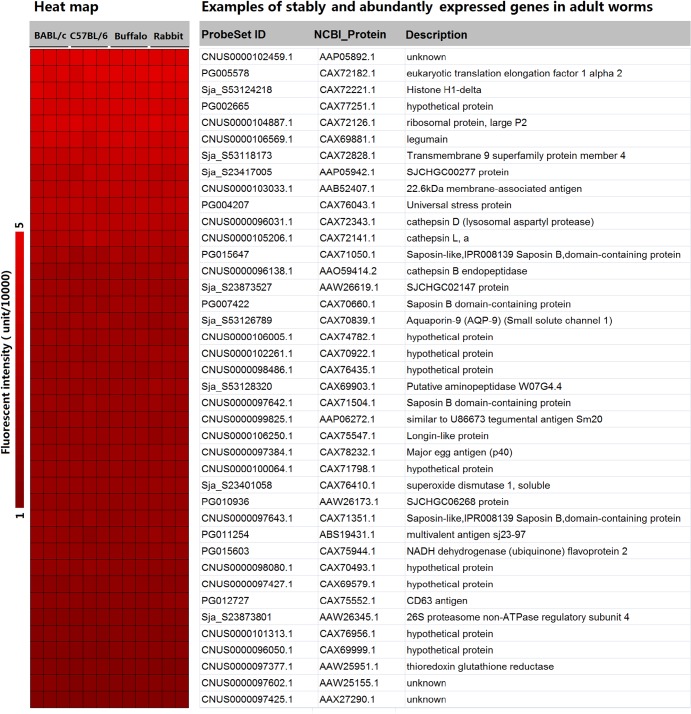
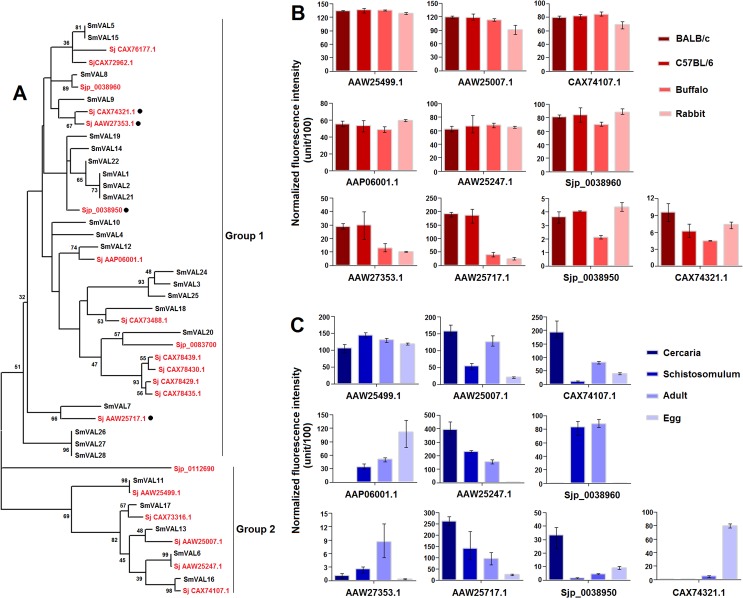
Methodology/principal findings: Adult schistosomes were obtained from laboratory animals BALB/c mice, C57BL/6 mice, New Zealand white rabbits and the natural host, water buffaloes. The gene expression profiles of schistosomes from these animals were obtained and compared by genome-wide oligonucleotide microarray analysis. The results revealed that the gene expression profiles of schistosomes from different laboratory animals and buffaloes were highly consistent (r>0.98) genome-wide. Meanwhile, a total of 450 genes were identified to be differentially expressed in schistosomes which can be clustered into six groups. Pathway analysis revealed that these genes were mainly involved in multiple signal transduction pathways, amino acid, energy, nucleotide and lipid metabolism. We also identified a group of 1,540 abundantly and stably expressed gene products in adult worms, including a panel of 179 Schistosoma- or Platyhelminthes-specific genes that may be essential for parasitism and may be regarded as novel potential anti-parasite intervention targets for future research.
Conclusions/significance: This study provides a comprehensive database of gene expression profiles of schistosomes derived from different laboratory animals and water buffaloes. An expanded number of genes potentially affecting the development of schistosomes in different animals were identified. These findings lay the foundation for schistosomiasis research in different laboratory animals and natural hosts at the transcriptional level and provide a valuable resource for screening anti-schistosomal intervention targets.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Steinmann P, Keiser J, Bos R, Tanner M, Utzinger J. Schistosomiasis and water resources development: systematic review, meta-analysis, and estimates of people at risk. Lancet Infect Dis. 2006;6(7):411–25. - PubMed
-
- Vos T, Flaxman AD, Naghavi M, Lozano R, Michaud C, Ezzati M, et al. Years lived with disability (YLDs) for 1160 sequelae of 289 diseases and injuries 1990–2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet. 2012;380(9859):2163–96. 10.1016/S0140-6736(12)61729-2 - DOI - PMC - PubMed
-
- Gryseels B, Polman K, Clerinx J, Kestens L. Human schistosomiasis. Lancet. 2006;368(9541):1106–18. - PubMed
-
- He YX, Salafsky B, Ramaswamy K. Host—parasite relationships of Schistosoma japonicum in mammalian hosts. Trends Parasitol. 2001;17(7):320–4. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases