Identifying genomic changes associated with insecticide resistance in the dengue mosquito Aedes aegypti by deep targeted sequencing
- PMID: 26206155
- PMCID: PMC4561493
- DOI: 10.1101/gr.189225.115
Identifying genomic changes associated with insecticide resistance in the dengue mosquito Aedes aegypti by deep targeted sequencing
Abstract
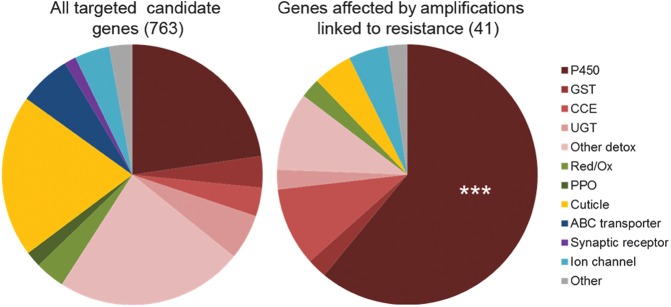
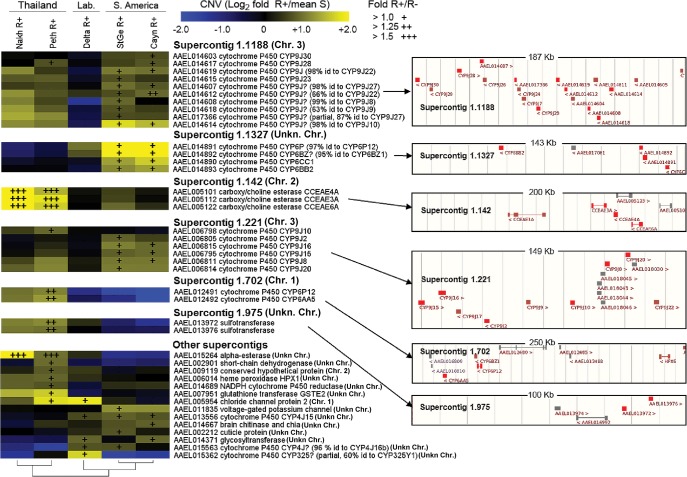
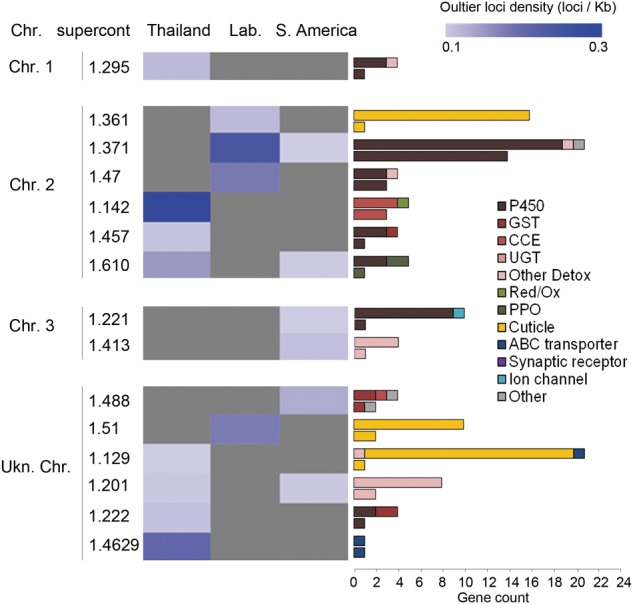
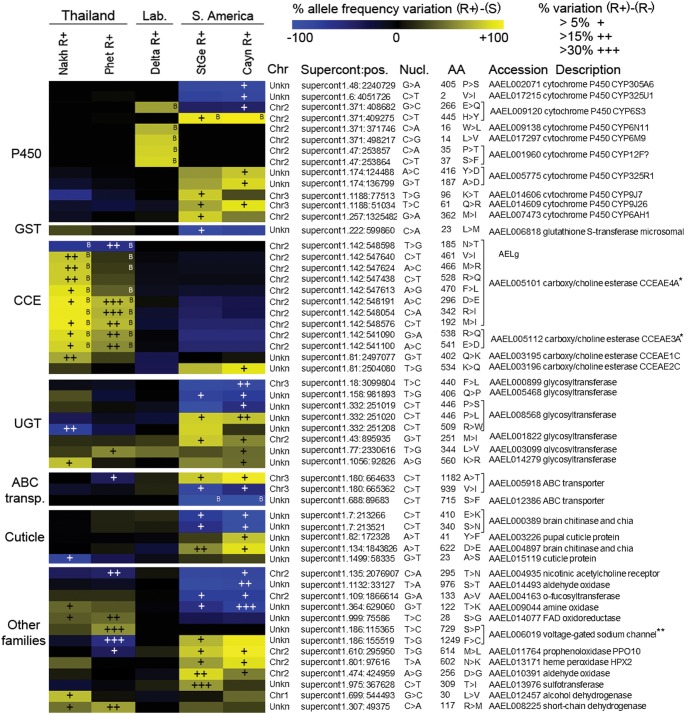
The capacity of mosquitoes to resist insecticides threatens the control of diseases such as dengue and malaria. Until alternative control tools are implemented, characterizing resistance mechanisms is crucial for managing resistance in natural populations. Insecticide biodegradation by detoxification enzymes is a common resistance mechanism; however, the genomic changes underlying this mechanism have rarely been identified, precluding individual resistance genotyping. In particular, the role of copy number variations (CNVs) and polymorphisms of detoxification enzymes have never been investigated at the genome level, although they can represent robust markers of metabolic resistance. In this context, we combined target enrichment with high-throughput sequencing for conducting the first comprehensive screening of gene amplifications and polymorphisms associated with insecticide resistance in mosquitoes. More than 760 candidate genes were captured and deep sequenced in several populations of the dengue mosquito Ae. aegypti displaying distinct genetic backgrounds and contrasted resistance levels to the insecticide deltamethrin. CNV analysis identified 41 gene amplifications associated with resistance, most affecting cytochrome P450s overtranscribed in resistant populations. Polymorphism analysis detected more than 30,000 variants and strong selection footprints in specific genomic regions. Combining Bayesian and allele frequency filtering approaches identified 55 nonsynonymous variants strongly associated with resistance. Both CNVs and polymorphisms were conserved within regions but differed across continents, confirming that genomic changes underlying metabolic resistance to insecticides are not universal. By identifying novel DNA markers of insecticide resistance, this study opens the way for tracking down metabolic changes developed by mosquitoes to resist insecticides within and among populations.
© 2015 Faucon et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Altmüller J, Budde BS, Nürnberg P. 2014. Enrichment of target sequences for next-generation sequencing applications in research and diagnostics. Biol Chem 395: 231–237. - PubMed
-
- Amichot M, Tarés S, Brun-Barale A, Arthaud L, Bride JM, Bergè JB. 2004. Point mutations associated with insecticide resistance in the Drosophila cytochrome P450 Cyp6a2 enable DDT metabolism. Eur J Biochem 1: 1250–1257. - PubMed
-
- Bass C, Field LM. 2011. Gene amplification and insecticide resistance. Pest Manag Sci 67: 886–890. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases