Pathogenic diversity amongst serotype C VGIII and VGIV Cryptococcus gattii isolates
- PMID: 26153364
- PMCID: PMC4495446
- DOI: 10.1038/srep11717
Pathogenic diversity amongst serotype C VGIII and VGIV Cryptococcus gattii isolates
Abstract
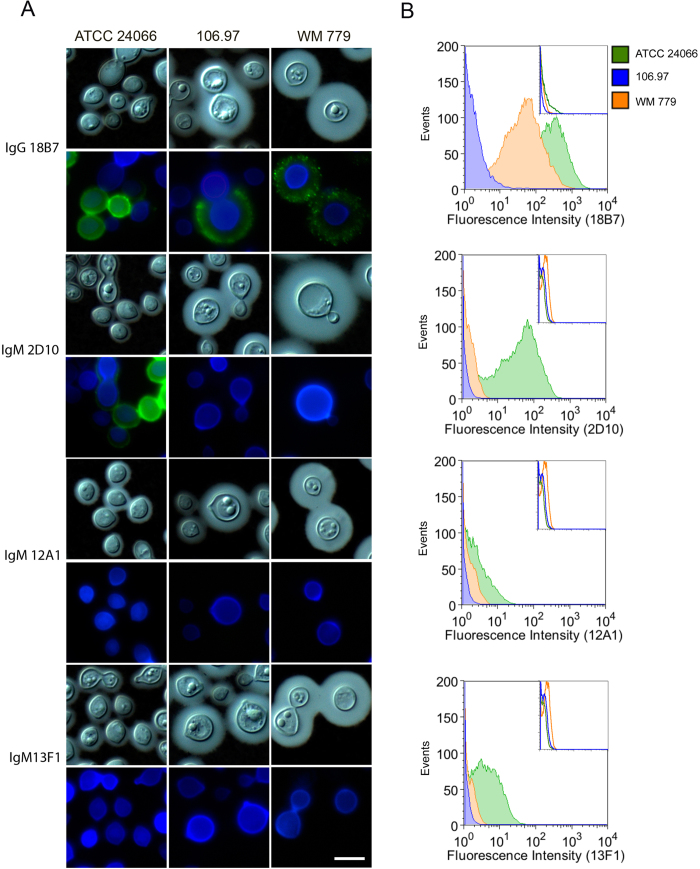
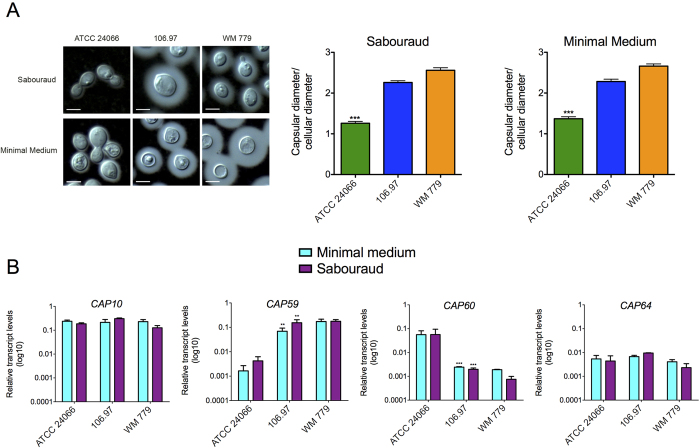
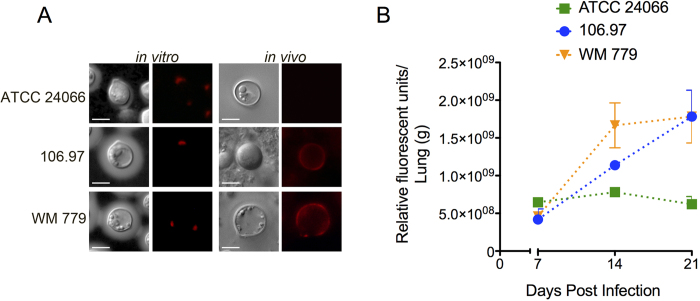
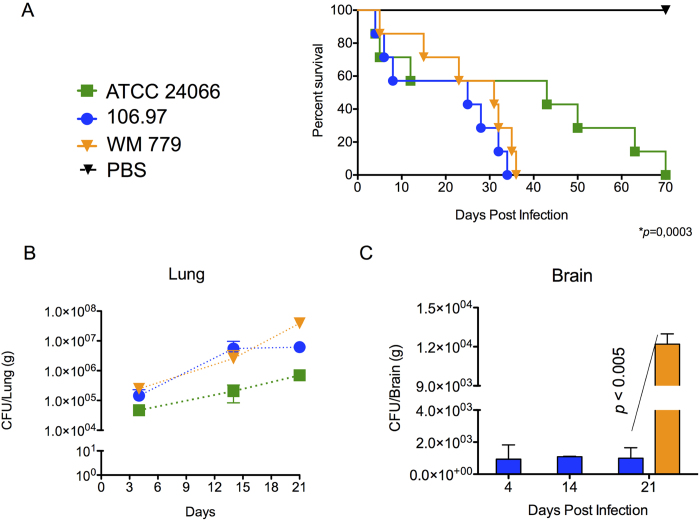
Cryptococcus gattii is one of the causative agents of human cryptococcosis. Highly virulent strains of serotype B C. gattii have been studied in detail, but little information is available on the pathogenic properties of serotype C isolates. In this study, we analyzed pathogenic determinants in three serotype C C. gattii isolates (106.97, ATCC 24066 and WM 779). Isolate ATCC 24066 (molecular type VGIII) differed from isolates WM 779 and 106.97 (both VGIV) in capsule dimensions, expression of CAP genes, chitooligomer distribution, and induction of host chitinase activity. Isolate WM 779 was more efficient than the others in producing pigments and all three isolates had distinct patterns of reactivity with antibodies to glucuronoxylomannan. This great phenotypic diversity reflected in differential pathogenicity. VGIV isolates WM 779 and 106.97 were similar in their ability to cause lethality and produced higher pulmonary fungal burden in a murine model of cryptococcosis, while isolate ATCC 24066 (VGIII) was unable to reach the brain and caused reduced lethality in intranasally infected mice. These results demonstrate a high diversity in the pathogenic potential of isolates of C. gattii belonging to the molecular types VGIII and VGIV.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Park B. J. et al. Estimation of the current global burden of cryptococcal meningitis among persons living with HIV/AIDS. AIDS 23, 525–30 (2009). - PubMed
-
- Chayakulkeeree M. & Perfect J. R. Cryptococcosis. Infect Dis Clin North Am 20, 507–44, v-vi (2006). - PubMed
-
- Huston S. M. & Mody C. H. Cryptococcosis: an emerging respiratory mycosis. Clin Chest Med 30, 253–64, vi (2009). - PubMed
-
- Lin X. Cryptococcus neoformans: morphogenesis, infection, and evolution. Infect Genet Evol 9, 401–16 (2009). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous