Complexity of Neutralizing Antibodies against Multiple Dengue Virus Serotypes after Heterotypic Immunization and Secondary Infection Revealed by In-Depth Analysis of Cross-Reactive Antibodies
- PMID: 25972550
- PMCID: PMC4473561
- DOI: 10.1128/JVI.00273-15
Complexity of Neutralizing Antibodies against Multiple Dengue Virus Serotypes after Heterotypic Immunization and Secondary Infection Revealed by In-Depth Analysis of Cross-Reactive Antibodies
Abstract
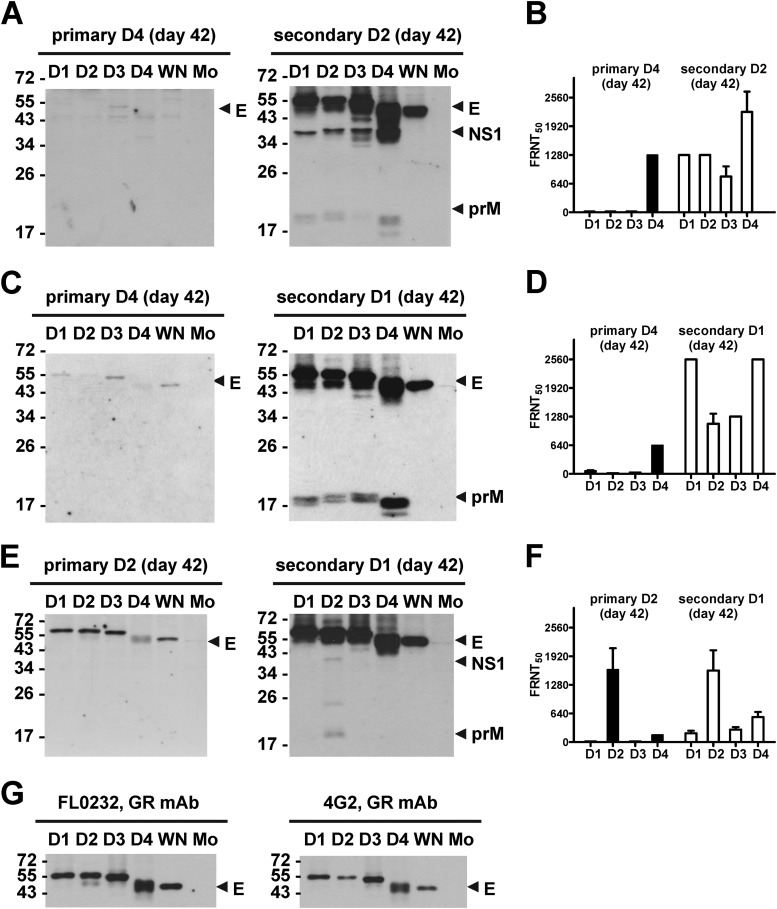
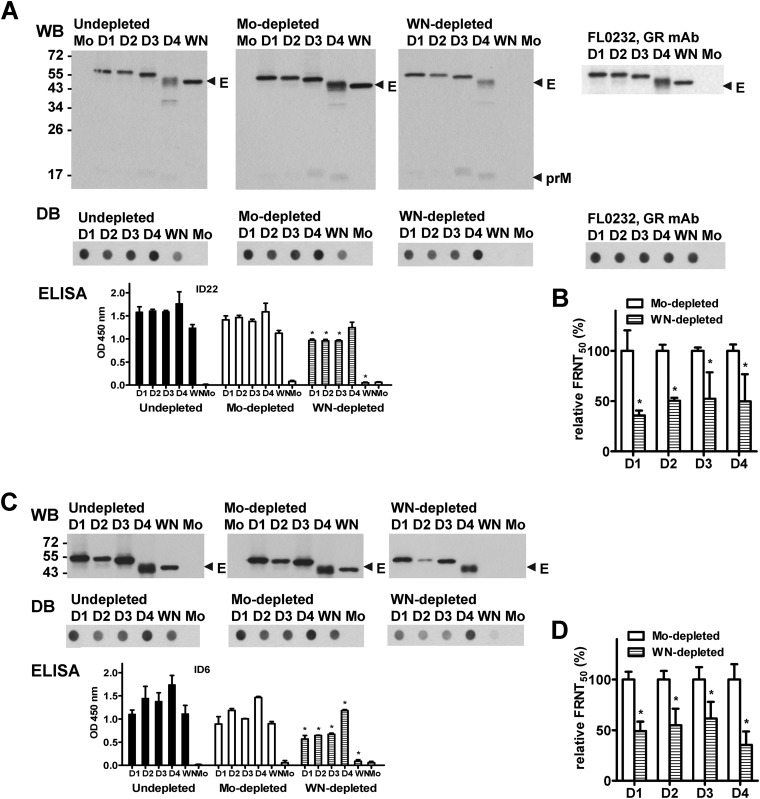
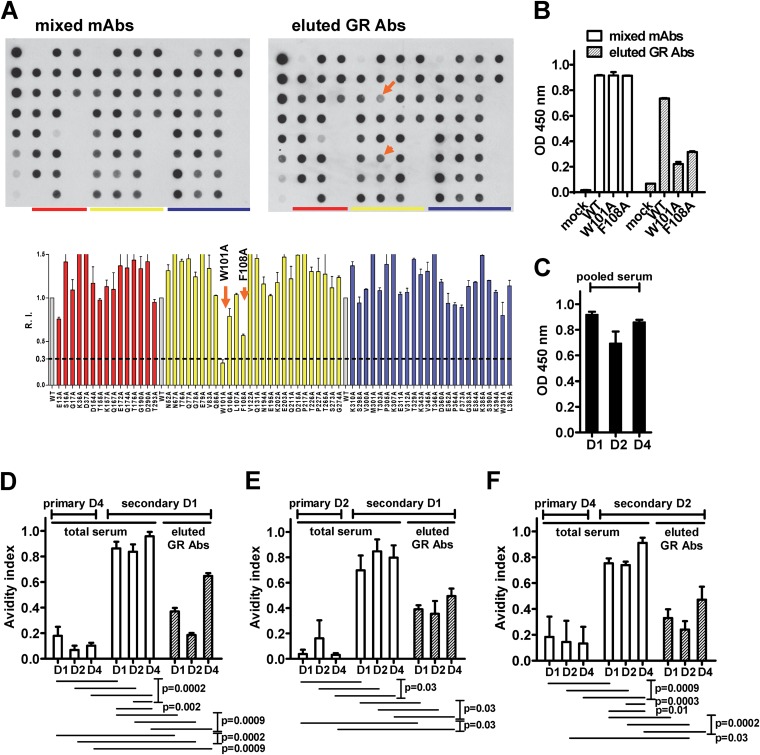
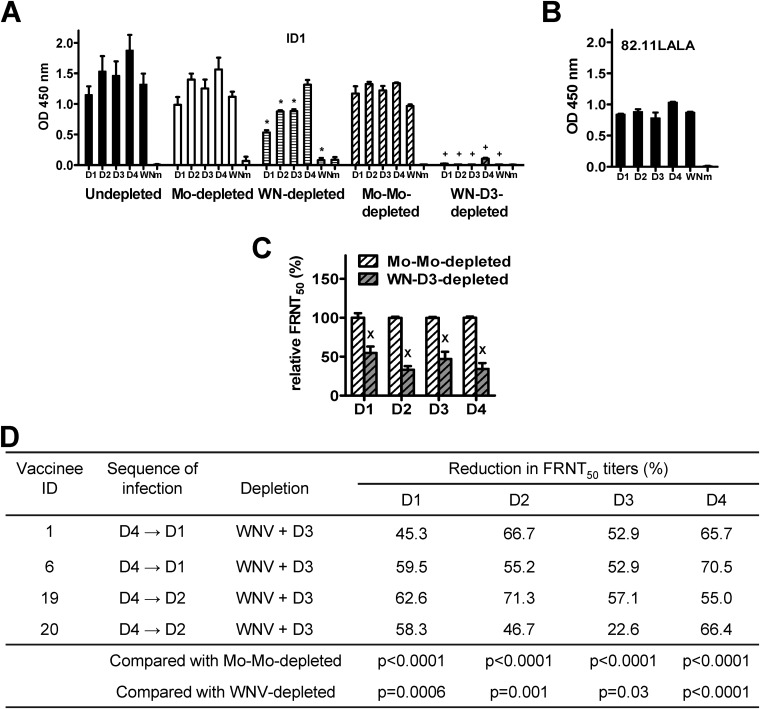
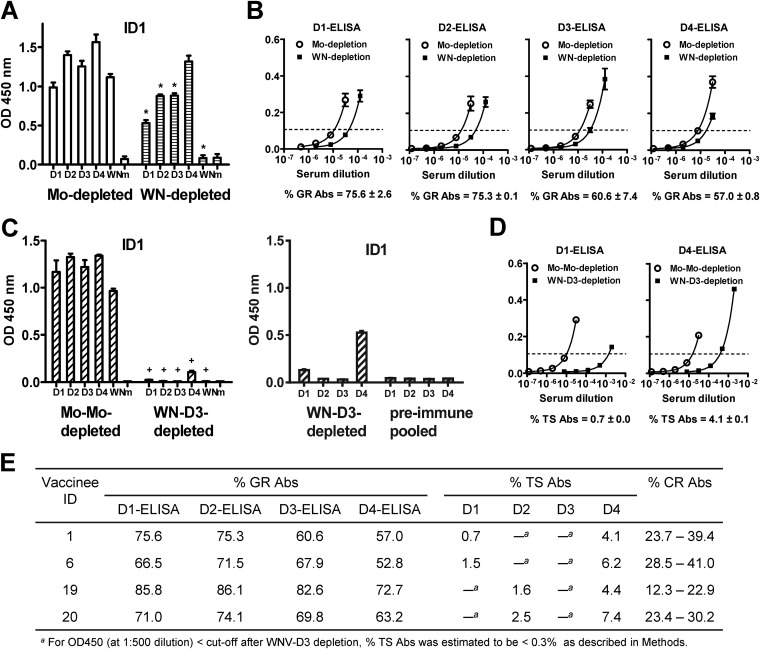
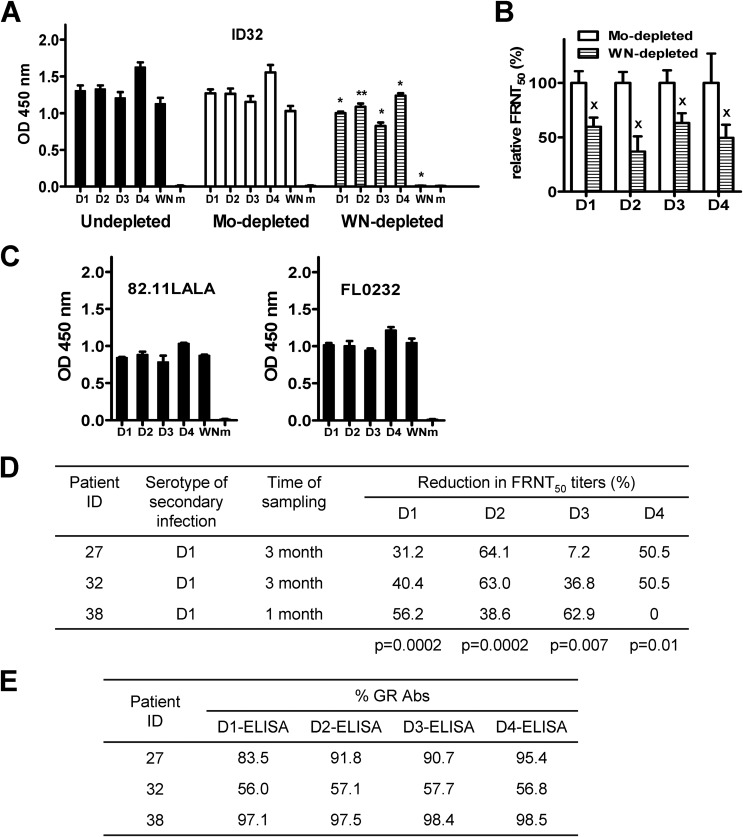
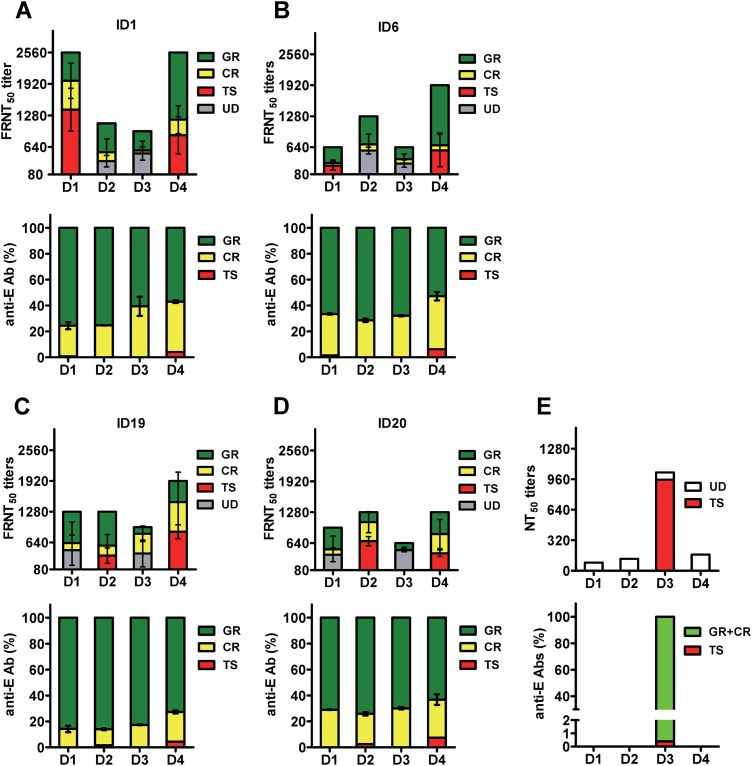
The four serotypes of dengue virus (DENV) cause the most important and rapidly emerging arboviral diseases in humans. The recent phase 2b and 3 studies of a tetravalent dengue vaccine reported a moderate efficacy despite the presence of neutralizing antibodies, highlighting the need for a better understanding of neutralizing antibodies in polyclonal human sera. Certain type-specific (TS) antibodies were recently discovered to account for the monotypic neutralizing activity and protection after primary DENV infection. The nature of neutralizing antibodies after secondary DENV infection remains largely unknown. In this study, we examined sera from 10 vaccinees with well-documented exposure to first and second DENV serotypes through heterotypic immunization with live-attenuated vaccines. Higher serum IgG avidities to both exposed and nonexposed serotypes were found after secondary immunization than after primary immunization. Using a two-step depletion protocol to remove different anti-envelope antibodies, including group-reactive (GR) and complex-reactive (CR) antibodies separately, we found GR and CR antibodies together contributed to more than 50% of neutralizing activities against multiple serotypes after secondary immunization. Similar findings were demonstrated in patients after secondary infection. Anti-envelope antibodies recognizing previously exposed serotypes consisted of a large proportion of GR antibodies, CR antibodies, and a small proportion of TS antibodies, whereas those recognizing nonexposed serotypes consisted of GRand CR antibodies. These findings have implications for sequential heterotypic immunization or primary immunization of DENV-primed individuals as alternative strategies for DENV vaccination. The complexity of neutralizing antibodies after secondary infection provides new insights into the difficulty of their application as surrogates of protection.
Importance: The four serotypes of dengue virus (DENV) are the leading cause of arboviral diseases in humans. Despite the presence of neutralizing antibodies, a moderate efficacy was recently reported in phase 2b and 3 trials of a dengue vaccine; a better understanding of neutralizing antibodies in polyclonal human sera is urgently needed.We studied vaccinees who received heterotypic immunization of live-attenuated vaccines, as they were known to have received the first and second DENV serotype exposures.We found anti-envelope antibodies consist of group-reactive (GR), complex-reactive (CR), and type-specific (TS) antibodies, and that both GR and CR antibodies contribute significantly to multitypic neutralizing activities after secondary DENV immunization. These findings have implications for alternative strategies for DENV vaccination. Certain TS antibodies were recently discovered to contribute to the monotypic neutralizing activity and protection after primary DENV infection; our findings of the complexity of neutralizing activities after secondary immunization/infection provide new insights for neutralizing antibodies as surrogates of protection.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Bhatt S, Gething PW, Brady OJ, Messina JP, Farlow AW, Moyes CL, Drake JM, Brownstein JS, Hoen AG, Sankoh O, Myers MF, George DB, Jaenisch T, Wint GR, Simmons CP, Scott TW, Farrar JJ, Hay SI. 2013. The global distribution and burden of dengue. Nature 496:504–507. doi:10.1038/nature12060. - DOI - PMC - PubMed
-
- World Health Organization. 2009. Dengue hemorrhagic fever: diagnosis, treatment, prevention and control, 3rd ed WHO, Geneva, Switzerland.
-
- Sabchareon A, Wallace D, Sirivichayakul C, Limkittikul K, Chanthavanich P, Suvannadabba S, Jiwariyavej V, Dulyachai W, Pengsaa K, Wartel TA, Moureau A, Saville M, Bouckenooghe A, Viviani S, Tornieporth NG, Lang J. 2012. Protective efficacy of the recombinant, live-attenuated, CYD tetravalent dengue vaccine in Thai schoolchildren: a randomised, controlled phase 2b trial. Lancet 380:1559–1567. doi:10.1016/S0140-6736(12)61428-7. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical