A review of Theileria diagnostics and epidemiology
- PMID: 25830110
- PMCID: PMC4356873
- DOI: 10.1016/j.ijppaw.201412006
A review of Theileria diagnostics and epidemiology
Abstract
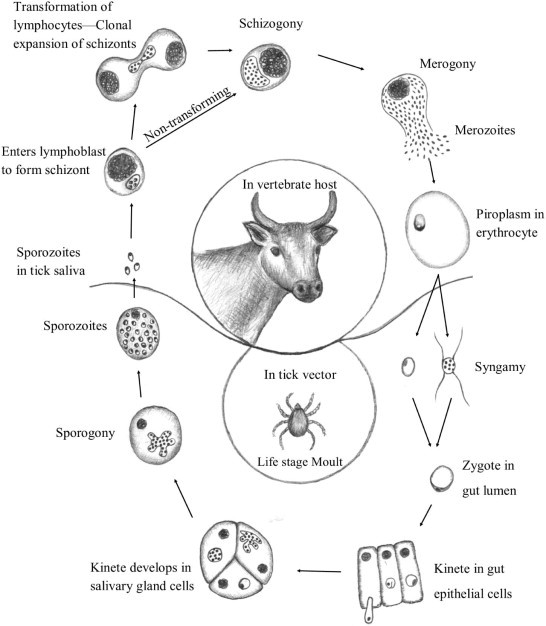
An extensive range of serological and molecular diagnostic assays exist for most of the economically important Theileira species such as T. annulata, T. equi, T. lestoquardi, T. parva, T. uilenbergi and other more benign species. Diagnostics of Theileria is considered with regard to sensitivity and specificity of current molecular and serological assays and their use in epidemiology. In the case of serological assays, cross-reactivity of genetically closely related species reduces the use of the gold standard indirect fluorescent antibody test (IFAT). Development of antigen-specific assays does not necessarily address this problem, since closely related species will potentially have similar antigens. Even so, serological assays remain an important line of enquiry in epidemiological surveys. Molecular based assays have exploded in the last decade with significant improvements in sensitivity and specificity. In this review, the current interpretation of what constitute a species in Theileria and its impact on accurate molecular diagnostics is considered. Most molecular assays based on conventional or real-time PCR technology have proven to be on standard with regard to analytical sensitivity. However, consideration of the limits of detection in regard to total blood volume of an animal indicates that most assays may only detect >400,000 parasites/L blood. Even so, natural parasitaemia distribution in carrier-state animals seems to be above this limit of detection, suggesting that most molecular assays should be able to detect the majority of infected individuals under endemic conditions. The potential for false-negative results can, however, only be assessed within the biological context of the parasite within its vertebrate host, i.e. parasitaemia range in the carrier-state that will support infection of the vector and subsequent transmission.
Keywords: Diagnostics; PCR; Parasitaemia; Serology; Species; Theileria.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Abdo J., Kristersson T., Seitzer U., Renneker S., Merza M., Ahmed J. Development and laboratory evaluation of a lateral flow device (LFD) for the serodiagnosis of Theileria annulata infection. Parasitol. Res. 2010;107:1241–1248. - PubMed
-
- Adili N., Melizi M. Preliminary study of the influence of red blood cells morphometry on the species determinism of domestic animals. Vet. World. 2014;7:219–223.
-
- Alhassan A., Pumidonming W., Okamura M., Hirata H., Battsetseg B., Fujisaki K. Development of a single-round and multiplex PCR method for the simultaneous detection of Babesia caballi and Babesia equi in horse blood. Vet. Parasitol. 2005;129:43–49. - PubMed
-
- Alhassan A., Govind Y., Tam N.T., Thekisoe O.M.M., Yokoyama N., Inoue N. Comparative evaluation of the sensitivity of LAMP, PCR and in vitro culture methods for the diagnosis of equine piroplasmosis. Parasitol. Res. 2007;100:1165–1168. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources