Predator diversity, intraguild predation, and indirect effects drive parasite transmission
- PMID: 25713379
- PMCID: PMC4364228
- DOI: 10.1073/pnas.1415971112
Predator diversity, intraguild predation, and indirect effects drive parasite transmission
Abstract
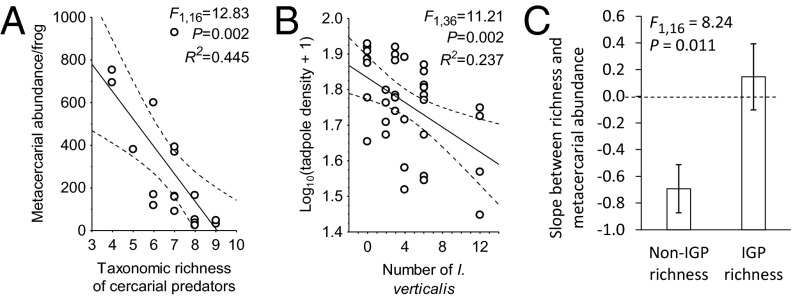
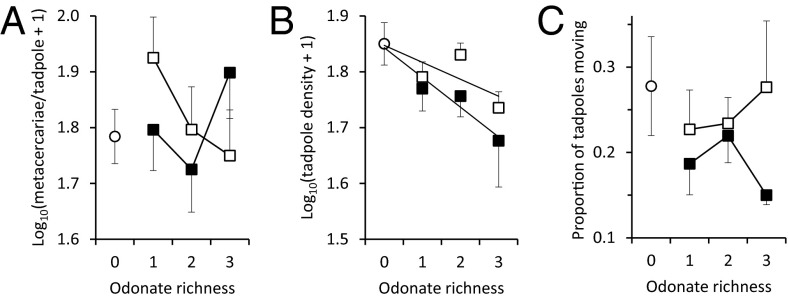
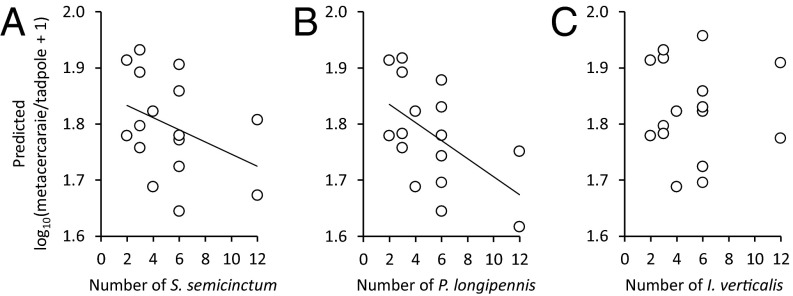
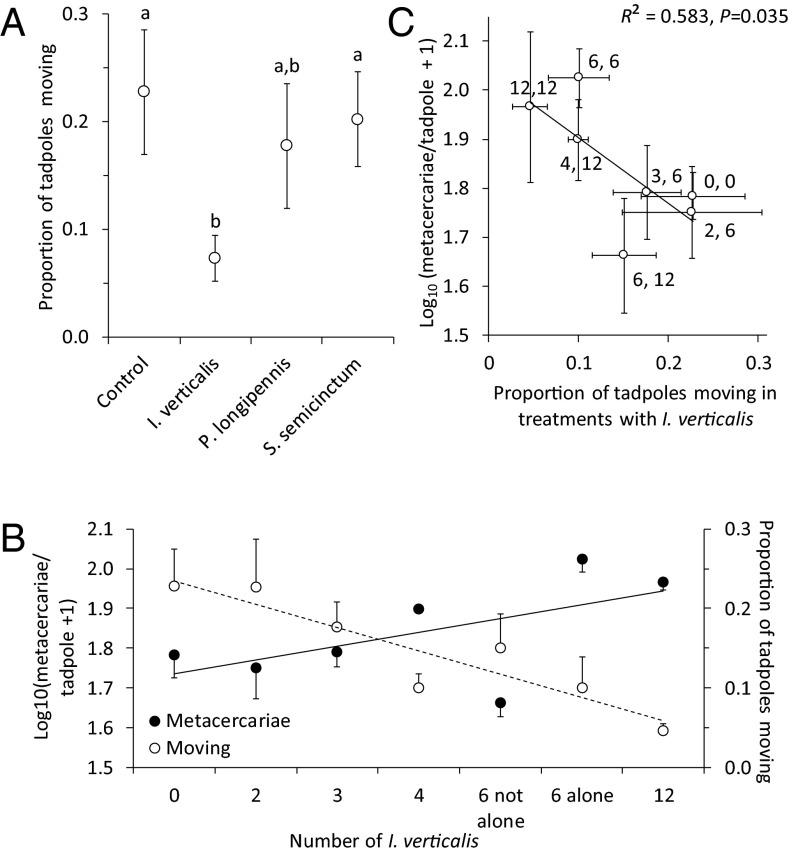
Humans are altering biodiversity globally and infectious diseases are on the rise; thus, there is interest in understanding how changes to biodiversity affect disease. Here, we explore how predator diversity shapes parasite transmission. In a mesocosm experiment that manipulated predator (larval dragonflies and damselflies) density and diversity, non-intraguild (non-IG) predators that only consume free-living cercariae (parasitic trematodes) reduced metacercarial infections in tadpoles, whereas intraguild (IG) predators that consume both parasites and tadpole hosts did not. This likely occurred because IG predators reduced tadpole densities and anticercarial behaviors, increasing per capita exposure rates of the surviving tadpoles (i.e., via density- and trait-mediated effects) despite the consumption of parasites. A mathematical model demonstrated that non-IG predators reduce macroparasite infections, but IG predation weakens this "dilution effect" and can even amplify parasite burdens. Consistent with the experiment and model, a wetland survey revealed that the diversity of IG predators was unrelated to metacercarial burdens in amphibians, but the diversity of non-IG predators was negatively correlated with infections. These results are strikingly similar to generalities that have emerged from the predator diversity-pest biocontrol literature, suggesting that there may be general mechanisms for pest control and that biocontrol research might inform disease management and vice versa. In summary, we identified a general trait of predators--where they fall on an IG predation continuum--that predicts their ability to reduce infections and possibly pests in general. Consequently, managing assemblages of predators represents an underused tool for the management of human and wildlife diseases and pest populations.
Keywords: biodiversity–ecosystem function; dilution effect; schistosomiasis; snail; trophic cascade.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials