Sm16, a major component of Schistosoma mansoni cercarial excretory/secretory products, prevents macrophage classical activation and delays antigen processing
- PMID: 25561160
- PMCID: PMC4297449
- DOI: 10.1186/s13071-014-0608-1
Sm16, a major component of Schistosoma mansoni cercarial excretory/secretory products, prevents macrophage classical activation and delays antigen processing
Abstract
Background: Schistosoma mansoni cercariae penetrate the skin by releasing excretory/secretory (E/S) products known as 0-3hRP, which are associated with immune modulation through Toll like receptor (TLR) signalling. Furthermore, these secretions contain Sm16, which when given to cells as a recombinant protein inhibits human monocyte derived cytokine responses to TLR4 and TLR3 ligands. Nonetheless, the extent and mechanism(s) of these inhibitory effects remain largely uncharacterized.
Methods: Murine bone marrow derived macrophages were exposed to different fractions of 0-3hRP, obtained via ultracentrifugation, or recombinant Sm16. These cells were exposed to the parasite molecules in combination with different TLR ligands, or Interferon gamma, and tested for the production of the cytokines IL-10 and IL-12p40, and their ability to process antigen.
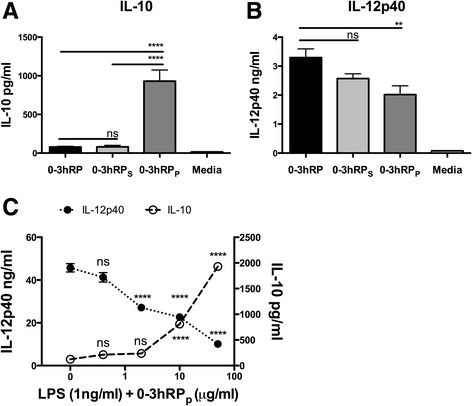
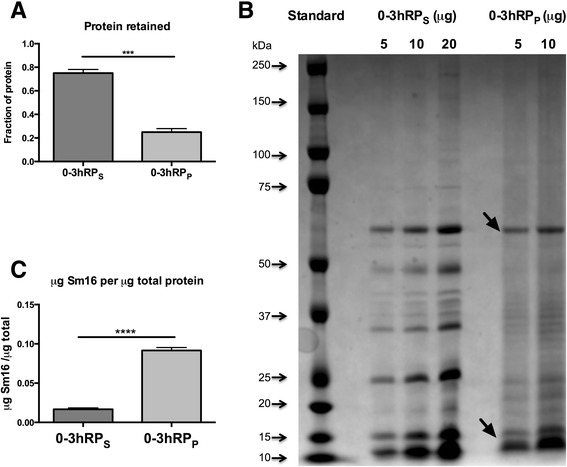
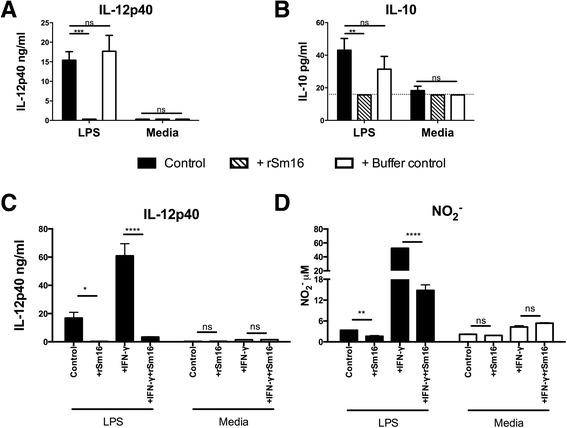
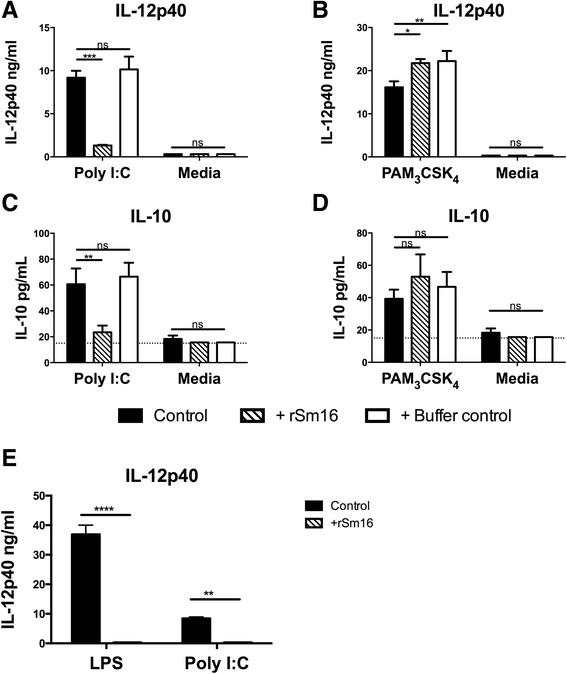
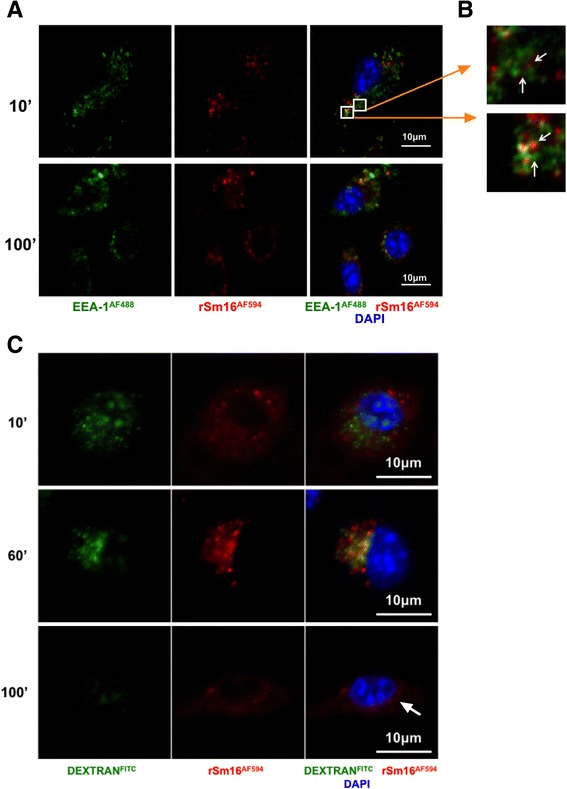
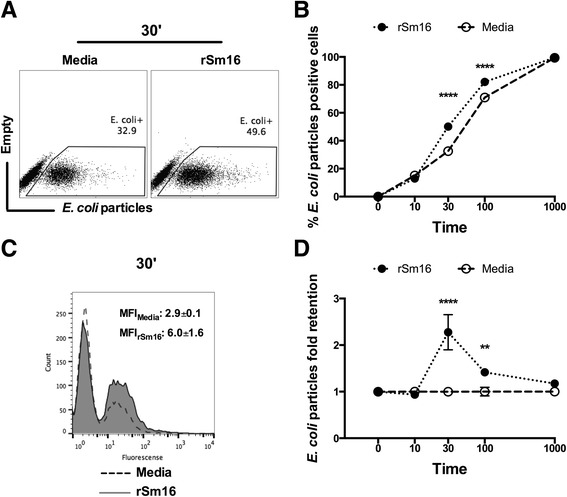
Results: The immunomodulatory function of 0-3hRP is enriched predominantly in the pellet fraction, which contains a greater proportion of Sm16, also corroborating the ability of recombinant Sm16 to inhibit macrophage activation in response to TLR ligands. We further demonstrate that Sm16 blocks classical activation of macrophages to LPS or IFN-γ stimulation in vitro, and that inhibition of macrophage classical activation is independent of TLR2 recognition. Finally we show that Sm16 shares the altered intracellular processing observed for 0-3hRP, and is able to delay antigen processing by macrophages.
Conclusions: Collectively, our findings show that Sm16 is a major component of S. mansoni cercarial E/S products, and is partly responsible for its immune-regulatory properties. Moreover, we propose that the mechanism employed by Sm16 to exert its inhibitory function is likely to be linked with alteration of endosomal trafficking and is not dependent on particular TLR receptors. Finally, we suggest that accumulation of Sm16 in the skin after percutaneous infection with S. mansoni cercariae could contribute to limiting dermal inflammation.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources