Properties of AdeABC and AdeIJK efflux systems of Acinetobacter baumannii compared with those of the AcrAB-TolC system of Escherichia coli
- PMID: 25246403
- PMCID: PMC4249520
- DOI: 10.1128/AAC.03728-14
Properties of AdeABC and AdeIJK efflux systems of Acinetobacter baumannii compared with those of the AcrAB-TolC system of Escherichia coli
Abstract
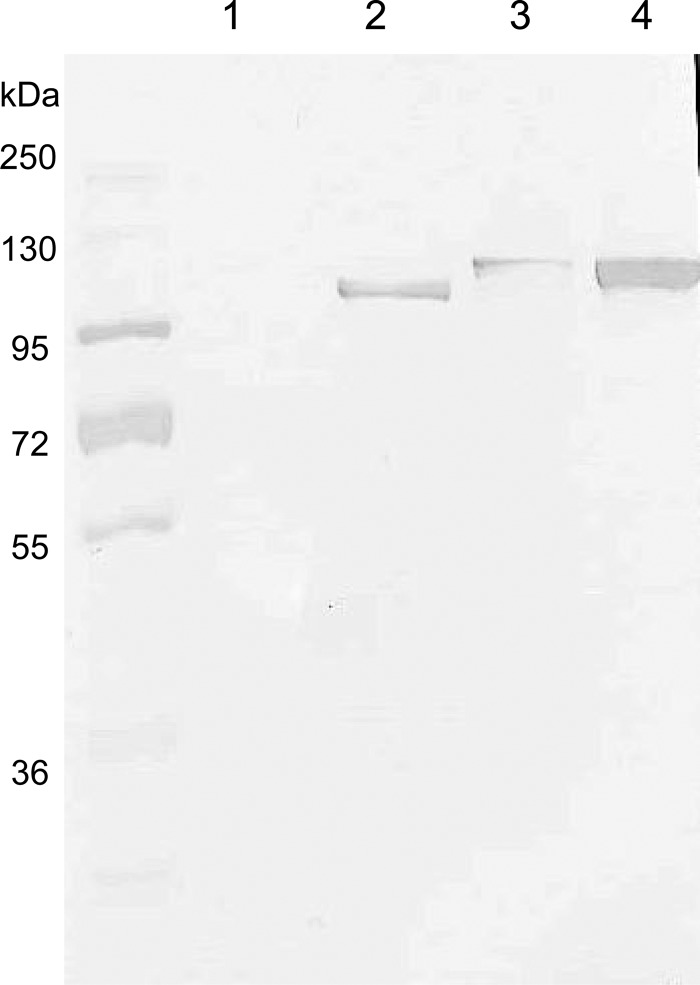
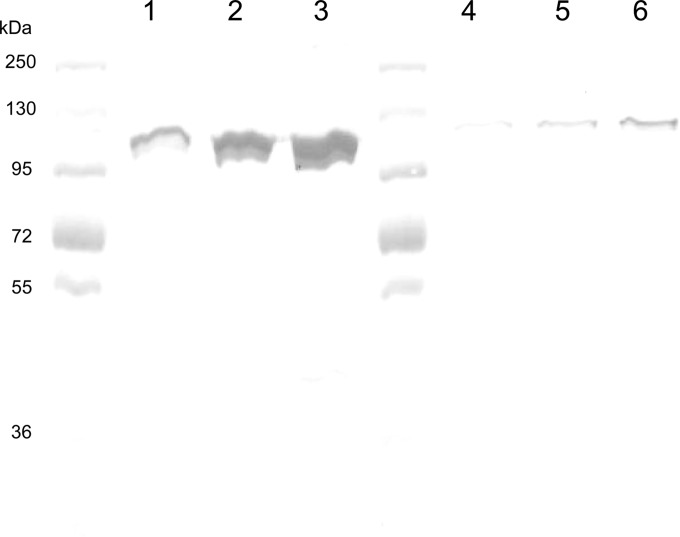
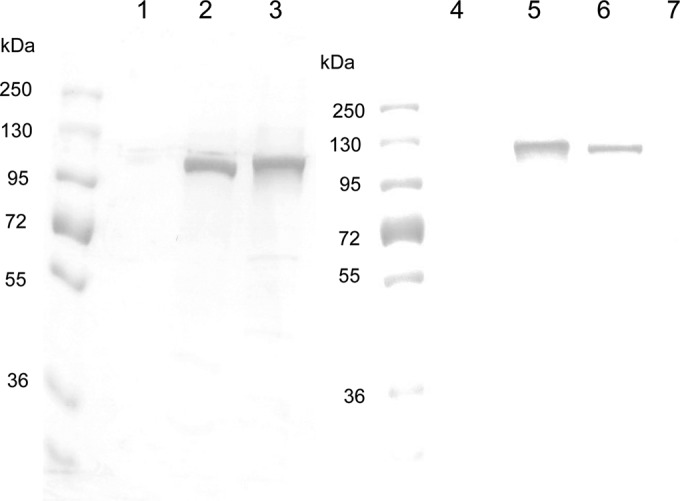
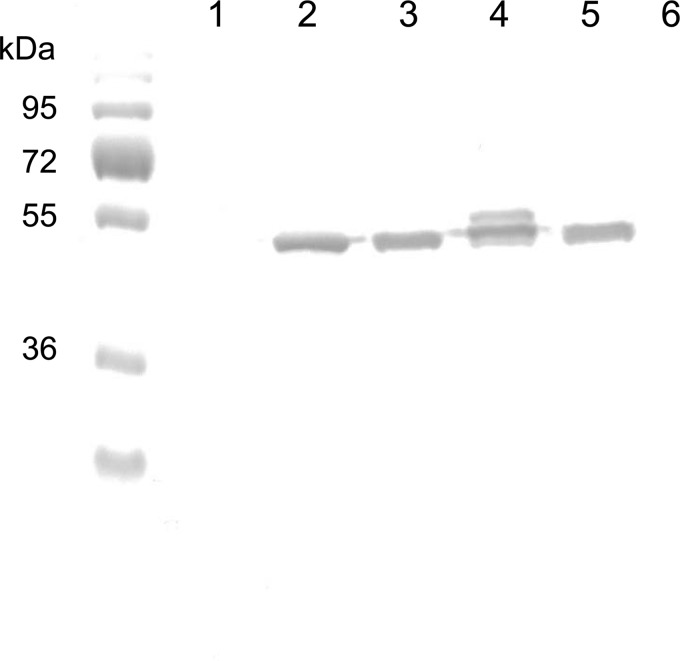
Acinetobacter baumannii contains RND-family efflux systems AdeABC and AdeIJK, which pump out a wide range of antimicrobial compounds, as judged from the MIC changes occurring upon deletion of the responsible genes. However, these studies may miss changes because of the high backgrounds generated by the remaining pumps and by β-lactamases, and it is unclear how the activities of these pumps compare quantitatively with those of the well-studied AcrAB-TolC system of Escherichia coli. We expressed adeABC and adeIJK of A. baumannii, as well as E. coli acrAB, in an E. coli host from which acrAB was deleted. The A. baumannii pumps were functional in E. coli, and the MIC changes that were observed largely confirmed the substrate range already reported, with important differences. Thus, the AdeABC system pumped out all β-lactams, an activity that was often missed in deletion studies. When the expression level of the pump genes was adjusted to a similar level for a comparison with AcrAB-TolC, we found that both A. baumannii efflux systems pumped out a wide range of compounds, but AdeABC was less effective than AcrAB-TolC in the extrusion of lipophilic β-lactams, novobiocin, and ethidium bromide, although it was more effective at tetracycline efflux. AdeIJK was remarkably more effective than a similar level of AcrAB-TolC in the efflux of β-lactams, novobiocin, and ethidium bromide, although it was less so in the efflux of erythromycin. These results thus allow us to compare these efflux systems on a quantitative basis, if we can assume that the heterologous systems are fully functional in the E. coli host.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Sugawara E, Nikaido H. 2004. OmpA/OprF: slow porins or channels produced by alternative folding of outer membrane proteins, p 119–138 In Benz R. (ed), Bacterial and eukaryotic porins: structure, function, mechanism. Wiley-VCH, Weinheim, Germany.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases