CFTR interacts with ZO-1 to regulate tight junction assembly and epithelial differentiation through the ZONAB pathway
- PMID: 25107366
- PMCID: PMC4197086
- DOI: 10.1242/jcs.148098
CFTR interacts with ZO-1 to regulate tight junction assembly and epithelial differentiation through the ZONAB pathway
Abstract
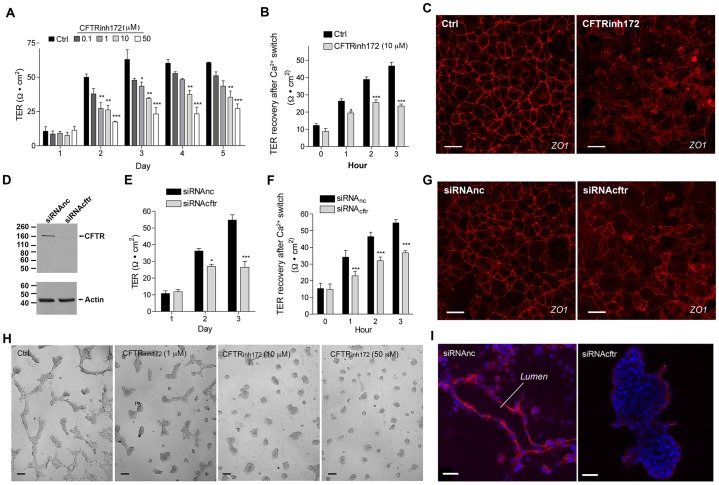
Mutations in CFTR lead to dysfunction of tubular organs, which is currently attributed to impairment of its conductive properties. We now show that CFTR regulates tight junction assembly and epithelial cell differentiation through modulation of the ZO-1-ZONAB pathway. CFTR colocalizes with ZO-1 at the tight junctions of trachea and epididymis, and is expressed before ZO-1 in Wolffian ducts. CFTR interacts with ZO-1 through the CTFR PDZ-binding domain. In a three-dimensional (3D) epithelial cell culture model, CFTR regulates tight junction assembly and is required for tubulogenesis. CFTR inhibition or knockdown reduces ZO-1 expression and induces the translocation of the transcription factor ZONAB (also known as YBX3) from tight junctions to the nucleus, followed by upregulation of the transcription of CCND1 and downregulation of ErbB2 transcription. The epididymal tubules of cftr(-/-) and cftr(ΔF508) mice have reduced ZO-1 levels, increased ZONAB nuclear expression, and decreased epithelial cell differentiation, illustrated by the reduced expression of apical AQP9 and V-ATPase. This study provides a new paradigm for the etiology of diseases associated with CFTR mutations, including cystic fibrosis.
Keywords: CFTR; Embryonic development; Epithelial remodeling; Male fertility; Morphogenesis; Proliferation; ZO-1; ZONAB.
© 2014. Published by The Company of Biologists Ltd.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Araki Y., Suzuki K., Matusik R. J., Obinata M., Orgebin-Crist M. C. (2002). Immortalized epididymal cell lines from transgenic mice overexpressing temperature-sensitive simian virus 40 large T-antigen gene. J. Androl. 23, 854–869 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous