Mechanisms of pyrethroid resistance in the dengue mosquito vector, Aedes aegypti: target site insensitivity, penetration, and metabolism
- PMID: 24945250
- PMCID: PMC4063723
- DOI: 10.1371/journal.pntd.0002948
Mechanisms of pyrethroid resistance in the dengue mosquito vector, Aedes aegypti: target site insensitivity, penetration, and metabolism
Abstract
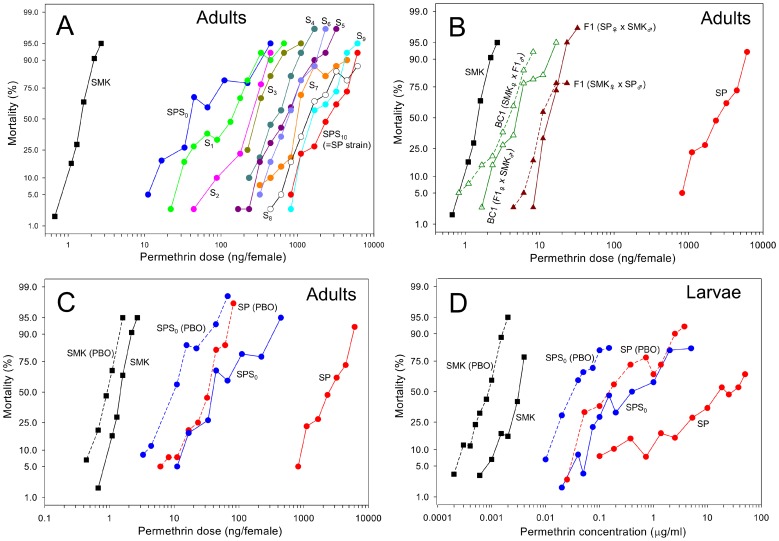
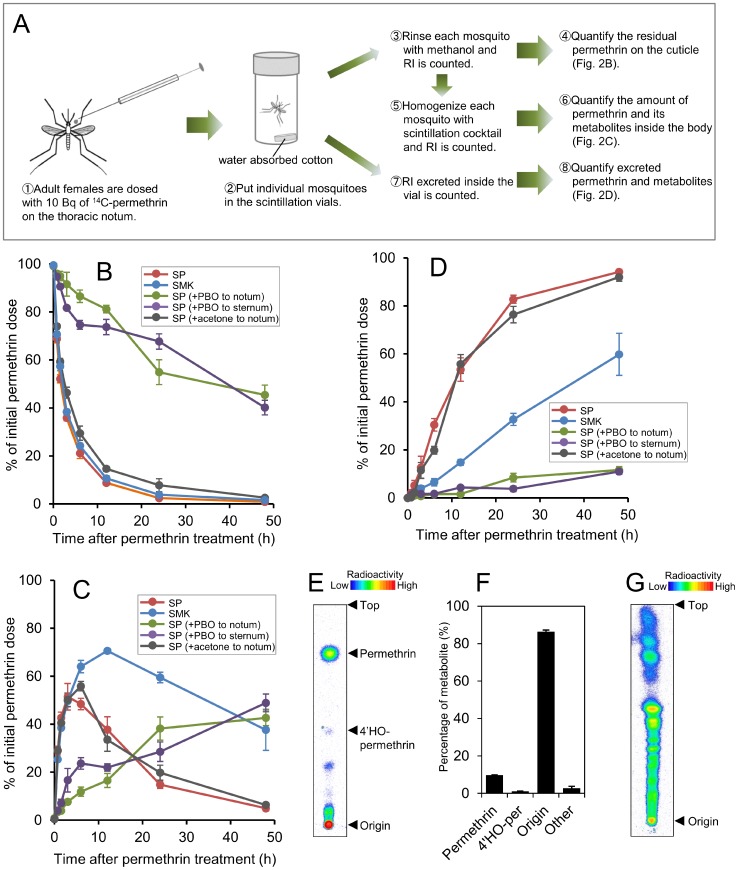
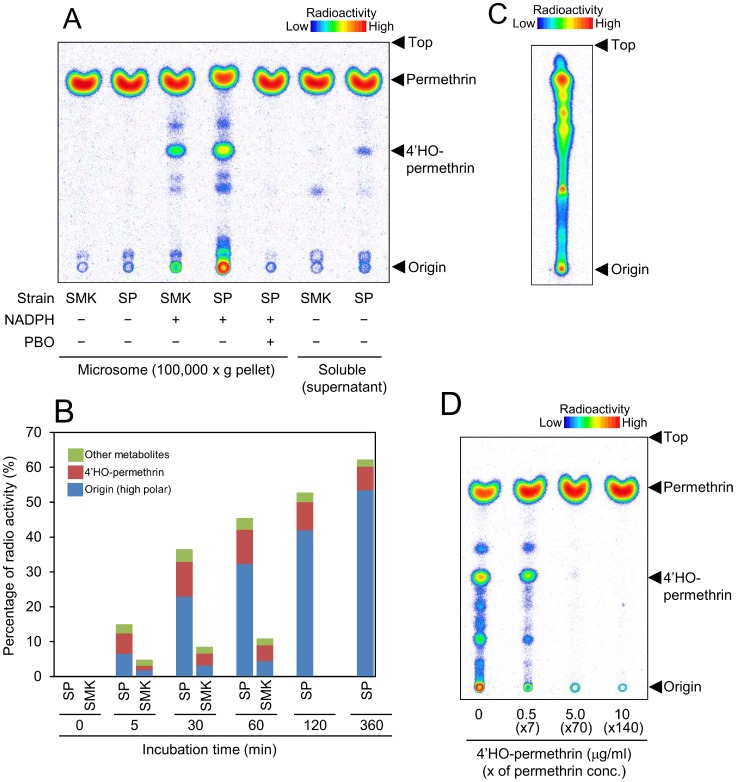
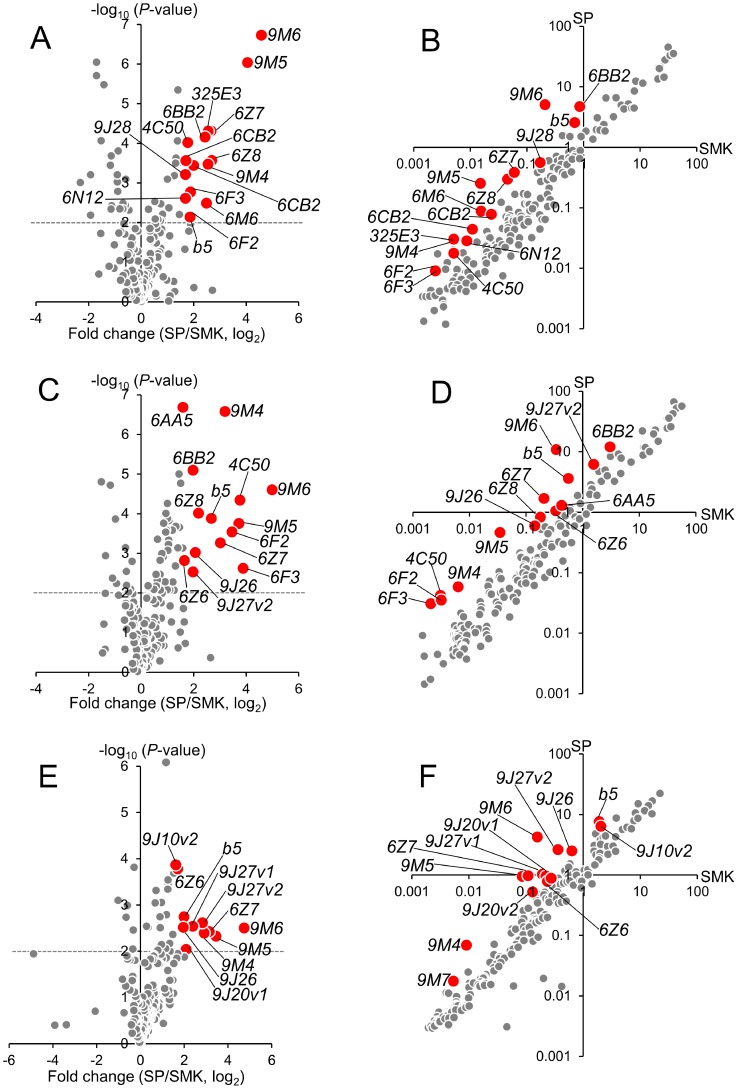
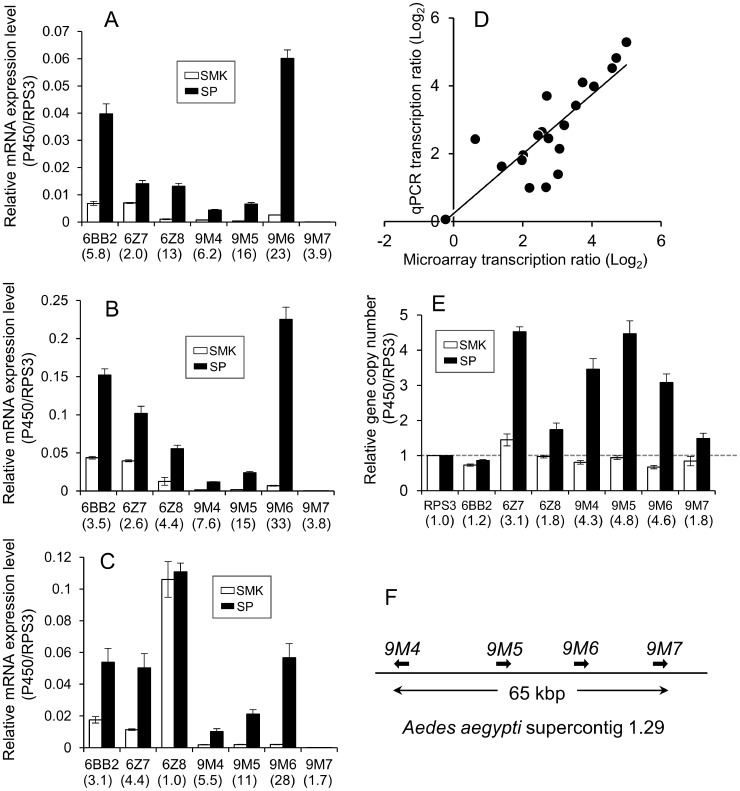
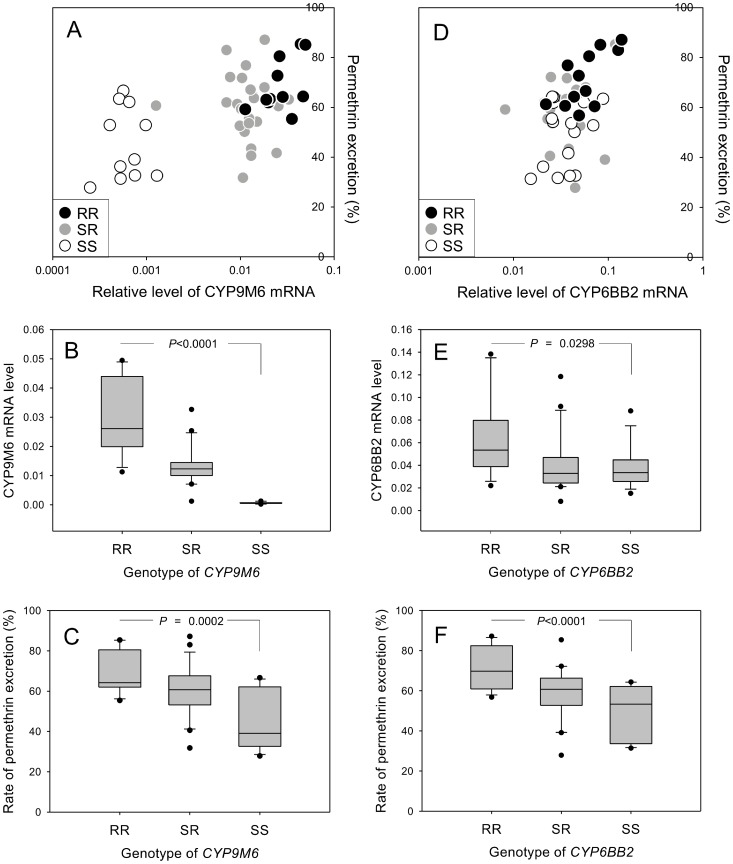
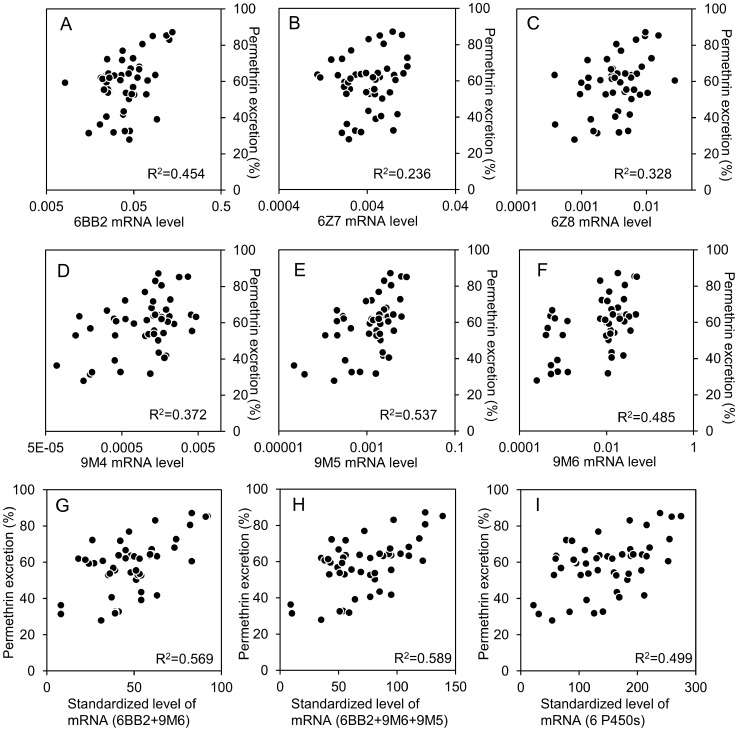
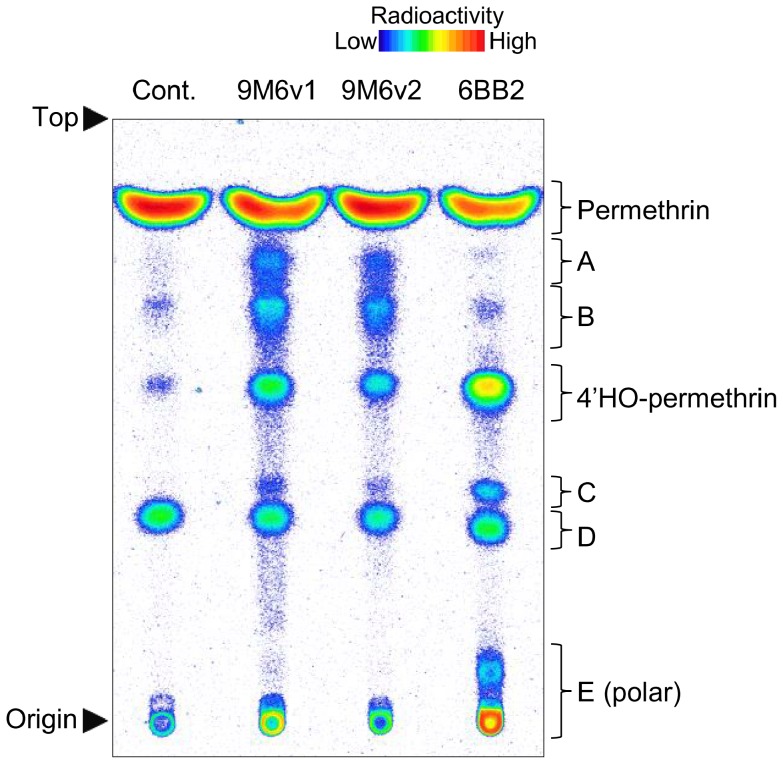
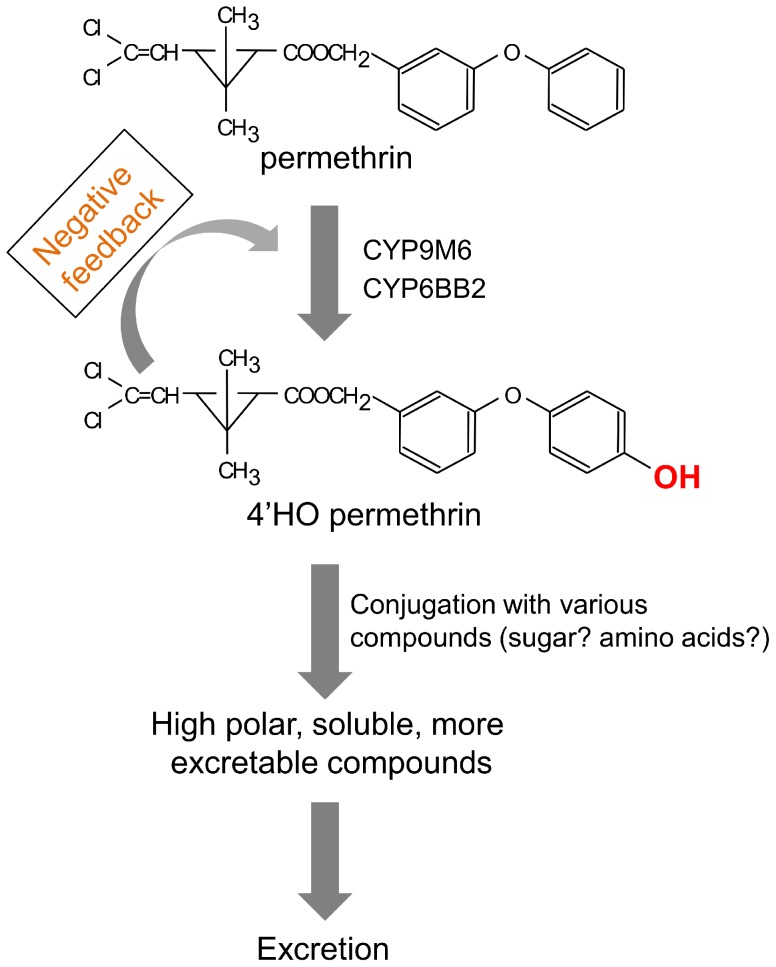
Aedes aegypti is the major vector of yellow and dengue fevers. After 10 generations of adult selection, an A. aegypti strain (SP) developed 1650-fold resistance to permethrin, which is one of the most widely used pyrethroid insecticides for mosquito control. SP larvae also developed 8790-fold resistance following selection of the adults. Prior to the selections, the frequencies of V1016G and F1534C mutations in domains II and III, respectively, of voltage-sensitive sodium channel (Vssc, the target site of pyrethroid insecticide) were 0.44 and 0.56, respectively. In contrast, only G1016 alleles were present after two permethrin selections, indicating that G1016 can more contribute to the insensitivity of Vssc than C1534. In vivo metabolism studies showed that the SP strain excreted permethrin metabolites more rapidly than a susceptible SMK strain. Pretreatment with piperonyl butoxide caused strong inhibition of excretion of permethrin metabolites, suggesting that cytochrome P450 monooxygenases (P450s) play an important role in resistance development. In vitro metabolism studies also indicated an association of P450s with resistance. Microarray analysis showed that multiple P450 genes were over expressed during the larval and adult stages in the SP strain. Following quantitative real time PCR, we focused on two P450 isoforms, CYP9M6 and CYP6BB2. Transcription levels of these P450s were well correlated with the rate of permethrin excretion and they were certainly capable of detoxifying permethrin to 4'-HO-permethrin. Over expression of CYP9M6 was partially due to gene amplification. There was no significant difference in the rate of permethrin reduction from cuticle between SP and SMK strains.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Mackenzie JS, Gubler DJ, Petersen LR (2004) Emerging flaviviruses: the spread and resurgence of Japanese encephalitis, West Nile and dengue viruses. Nat Med 10: S98–S109. - PubMed
-
- Gubler DJ (2002) Epidemic dengue/dengue hemorrhagic fever as a public health, social and economic problem in the 21st century. Trends Microbiol 10: 100–103. - PubMed
-
- Whitehorn J, Farrar J (2010) Dengue. Br Med Bull 95: 161–173. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources